Parte III: A biologia da reprodução e o hibridismo
A — Polimorfismos intraspecíficos
Dois polimorfismos intraspecíficos (locus Pgm de azinheira e locus Pgi-B de sobreiro) foram analisados em detalhe para caracterizar a biologia de reprodução das duas espécies e assim verificar se se aplicam (ou não) certos postulados relativos à incidência de híbridos. Tanto quanto é dado verificar, o de Pgm não foi descrito em azinheiras em nenhum outro estudo até à data.
Sendo inabitual recorrer-se a um único locus por espécie para tirar quaisquer conclusões, não deixa de ser verdade que os resultados da análise nesse locus, se for neutral e a amostragem das populações representativa e com dimensão suficiente, são teoricamente os mesmos que para todos os segmentos do genoma que também forem neutrais.
Tanto o locus Pgm de azinheira como o Pgi-B de sobreiro sugerem que os povoamentos mistos eram suficientemente homogéneos entre si para serem considerados (em cada espécie) uma só população. Podem considerar-se pelo menos dois factores, eventualmente convergentes entre si, para este resultado: primeiro, nuvens polínicas comuns (a partir das frequências dos gâmetas masculinos das tabelas 4.30 e 4.31, determina-se um GST de 2,21% para Pgm, relativamente baixo, e de 0,51% para Pgi-B, ainda mais baixo), sugerindo que a migração via pólen é suficientemente intensa para contribuir para a homogeneidade, apesar das distâncias serem na ordem das dezenas de quilómetros; e segundo, a intervenção de forças selectivas homogenizadoras, por exemplo selecção contra os homozigóticos (cf. secção 2).
Quando testada a hipótese das frequências do conjunto dos povoamentos mistos (adultos) serem próximas das previstas pelo modelo de Hardy-Weinberg, ela não foi rejeitada nem para Pgm nas azinheiras nem para Pgi-B nos sobreiros. Já nas populações de referência (plantas de viveiro) nota-se o contraste entre os dados para Pgm (desvios quando muito pouco significativos) e os de Pgi-B (rejeição da hipótese com erro tipo I de 1,2 × 10–5), neste último devido ao pronunciado déficit de heterozigóticos “ab” (e concomitante excesso de homozigóticos “a” e “b”), traduzido num coeficiente de fixação F = 0,33. Estes resultados sugerem duas coisas: primeiro, que a estrutura populacional varia com a fase do ciclo de vida; segundo, que as condicionantes sobre os dois loci em causa são diferentes — parece mais apropriado referirem-se estas diferenças aos loci e não às espécies, ou seja, implicar mecanismos selectivos específicos para cada locus em vez de postular diferenças na biologia populacional de sobreiro e azinheira, que parecem ser mínimas. Esta questão também será retomada na secção 2.
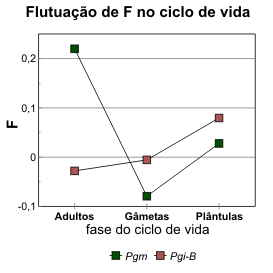
Uma maneira de definir um coeficiente de fixação é como uma medida, positiva ou negativa, do afastamento da frequência de heterozigóticos observados (Ho) em relação à esperada (He) no modelo de Hardy-Weinberg. Dispondo de estimativas de F para adultos, gâmetas e plantas jovens nos povoamentos mistos, observam-se flutuações neste parâmetro que, salvo erros de amostragem, podem ser interpretados em função de factores populacionais que possam afectar a frequência de heterozigóticos. Na figura 5.3 representam-se estas flutuações para os loci Pgm de azinheira e Pgi-B de sobreiro.
|
|
As estimativas de F na fase gamética são os valores de Fesp obtidos no modelo II (tabela 4.32), isto é, são a expressão da correlação negativa entre as frequências alélicas nos gâmetas que originaram as plantas analisadas, estimada a partir da diferença entre as frequências no pólen e as frequências nas plantas (cf. Apêndice II secção C). Esta correlação negativa é particularmente acentuada no locus Pgm e parece significar uma forte selecção contra a formação de homozigóticos na fase pré-zigótica, entre a polinização e a fertilização. Em contraste, no locus Pgi-B quase não há desvio de Fesp em relação ao valor 0 (o que se reflecte na semelhança das estimativas de Ne pelos dois modelos, na tabela 4.32).
Considerando as condições ideais de armazenamento, sementeira e criação em viveiro, e
assumindo que nenhum destes loci está envolvido em variações na capacidade de germinação das
sementes, é plausível que as flutuações de F desde a conjugação até às plantas de viveiro são neutrais. Se
o forem, então o ΔF na tabela 4.32 será uma boa estimativa do acréscimo de homozigóticos na
conjugação que reflecte a limitada diversidade genética disponível para fertilizar cada árvore, como
indicam os valores de Ne na tabela 4.32 (4,66 para Pgm e 5,87 para Pgi-B). O gráfico da figura 5.3
mostra como o aumento do F nesta fase é pronunciado, e quase paralelo entre os dois loci, como aliás
se esperaria entre espécies com biologias de reprodução semelhantes
![]() .
.
Se a taxa de autopolinização for baixa, como parece poder generalizar-se neste género [Cecich 1997a, Yacine & Bouras 1997, Boavida et al. 2001, Williams et al. 2001, Fernández-M. & Sork 2005], um valor de Ne baixo implica que o pólen aparentado geneticamente terá de vir doutras árvores, nomeadamente daquelas (poucas) na imediata vizinhança que estarão individualmente mais representadas na descendência (cf. “Revisão bibliográfica” parte II secção D1). Admitindo que nos adultos férteis não existe autocorrelação espacial, e portanto que não há parentesco entre os polinizadores mais próximos e as árvores polinizadas, os baixos valores de Ne terão de explicar-se por factores como a dispersão intraspecífica das datas de ântese desses polinizadores (cf. parte II, secção C1; efeito de Wahlund “temporal” [Chung et al. 2002]).
Tendo em conta a estimativa de FST para Pgm nas plantas jovens, e consequente dedução duma estimativa de Nem assumindo neutralidade selectiva, a utilização do valor Ne = 4,66 no mesmo material implica um valor bastante elevado da taxa de migração m = 0,315. Em princípio, este valor deve interpretar-se como um indicador de grande fluxo genético entre os povoamentos, o que pode admitir-se no abstracto, tendo em conta as observações que a maior parte do pólen que fertiliza cada árvore provém de fora do respectivo povoamento [Streiff et al. 1999]. Mas a distância entre os povoamentos mistos está para além do alcance médio das nuvens polínicas, estimado em menos de 10 Km noutros Quercus [Streiff et al. 1999].
Uma explicação alternativa para o valor de m encontrado, e que permanece válida mesmo com a estratificação dentro de cada povoamento (baixo Ne), é a existência de selecção estabilizante que faça convergir as frequências dos diferentes povoamentos para valores semelhantes (cf. “Revisão bibliográfica” parte IV, secção B1). Como ilustra a figura 4.17, as diferenças em Pgm entre as frequências nos gâmetas foram muito pronunciadas (daí a correlação negativa expressa em Fesp), sugerindo uma selecção pré-zigótica análoga à autoincompatibilidade, que pode considerar-se do tipo estabilizante. No caso de Pgi-B não existe nenhuma evidência directa da acção selectiva, a não ser que se assuma que o valor de F voltaria a ser negativo depois do desenvolvimento dos indivíduos que sobrevivessem até à fase adulta no local definitivo, por sobrevivência preferencial dos heterozigóticos neste locus. As actividades PGI são notórias pela tendência para a complementaridade funcional entre isoenzimas que favorece os heterozigóticos, fazendo recair a acção da selecção estabilizante no próprio locus e não noutros loci da mesma região cromossómica [Watt 1983, 1992]. A melhor maneira de colocar à prova esta hipótese será a verificação da sua generalidade (isto é, a variação do gráfico na figura 5.3) em tantos povoamentos quantos seja possível, tanto nos sobreiros como também em azinheiras. E, evidentemente, com maior número de famílias por povoamento.
Os valores de F nas plantas das populações de referência são superiores aos respectivos dos povoamentos mistos (particularmente na de sobreiro, F = 0,33), mas é possível que a maior estratificação aparente nessas populações reflicta apenas o modo como foram colhidos os lotes de semente; no essencial, não há contradição com os resultados nas plantas na figura 5.3, pois reitera-se o valor mais elevado de Pgi-B em relação ao de Pgm.
O modelo II foi importante sobretudo para corrigir a análise no locus Pgm, onde a correlação genética negativa entre os gâmetas foi mais acentuada (Fesp = –0,079). A escolha da diferença entre as frequências no pólen (estimadas por análise familiar) e nas plantas jovens, para determinação dos Δxi, resulta de serem as que provavelmente têm menor erro de amostragem, dada a dimensão da mesma (304 indivíduos). O valor de F neste locus, para os adultos (figura 5.3), é provavelmente um reflexo da amostragem limitada (60 indivíduos).
A possibilidade de haver contaminantes entre famílias, não detectados, é uma fonte de erro nestas frequências que seria evitada se se assegurasse que a colheita das sementes era feita directamente na árvore.
B — Implicações para a formação de híbridos
Na secção anterior deduziram-se aspectos da biologia da reprodução de azinheiras e sobreiros que podem considerar-se também factores relevantes para a formação de híbridos entre estas duas espécies.
Considerando que a possibilidade de ocorrer um híbrido implica uma exposição substancial duma das espécies ao pólen da outra espécie, e que os indivíduos mais próximos topograficamente e com data de floração próxima são os mais representados na descendência, então o risco de hibridismo será mais previsível se se verificarem cumulativamente:
i) sobreiros precoces na imediata vizinhança de azinheiras tardias,
ii) pólen daqueles a atingir estigmas receptivos destas (eventualmente vice-versa), e
iii) suficiente compatibilidade entre indivíduos.
A condição ii dependerá fortemente das datas de floração em cada ano, sendo os anos com um período de tempo significativo de sobreposição entre as florações das duas espécies os que requerem maior atenção. As condições i e iii são relativas ao grau de risco de hibridismo e justificariam um estudo especializado nos casos que correspondam à condição i. No caso dos três povoamentos mistos estudados, há muitos contactos próximos entre indivíduos das duas espécies (cf. Apêndice I secção B), mas é possível que em nenhum caso houvesse simultaneidade de floração entre árvores contíguas (pelo menos na Mitra isso é certo para 1998 [Varela et al. em preparação]).
A existência de selecção estabilizante, sugerida no presente trabalho tanto para o locus Pgm de azinheira (correlação negativa entre gâmetas) como para o Pgi-B de sobreiro (eliminação preferencial de plantas homozigóticas), não deve generalizar-se a outros loci que em cada espécie são monomórficos e nas linhagens híbridas passam a ser heterozigóticos. Por recombinação nos híbridos, dá-se início a novas combinações genotípicas interespecíficas que normalmente são inconsistentes adaptativamente (quando não o sejam, dá-se a oportunidade para surgir uma nova hibri-espécie), que quando muito tenderão a reconstituir as espécies originais por retrocruzamento (cf. parte II secção A2).
A amostragem de materiais orientada por estas hipóteses sobre os riscos de hibridação, desde que se aumente a informação sobre a sua plausibilidade, pode vir a ser uma abordagem complementar à do rastreio de proveniências (cf. parte II, secção D1), e a obrigatoriedade de acompanhar o progresso nesta linha de análise com um estudo mais aprofundado da biologia de reprodução intra-específica, se possível com um grande número de loci, transcende até a própria temática do hibridismo, embora lhe seja interdependente.