Parte II: Baixa probabilidade de formação de híbridos
Os carvalhos cerqueiros são bem conhecidos das gentes do Alentejo, e todos os que são referenciados no presente trabalho foram indicados por técnicos, guardas ou particulares. Dos 9 caracterizados com marcadores isoenzimáticos, todos apresentaram fenótipos híbridos, e somente SM2 talvez não seja híbrido de primeira geração mas sim descendente de híbrido (de SM1?, cf. parte I secção A). Ocorrem todos na proximidade de sobreiros e azinheiras, mas a sua distribuição geográfica parece preferencial para certos locais (figura 5.1): só em S. Marcos da Abóbada/Outeiro foram encontrados quatro (cf. Apêndice I, secção A2), e os híbridos CAB, VR e ALC distam poucos quilómetros entre si. Se se acrescentar a localização do presumível híbrido de Testos (cf. Apêndices I, secção A2, e II, secção D) e dos referenciados próximo do VR e na extrema da propriedade de Feijoas do Ramos, observa-se uma distribuição geográfica que, apesar dos poucos indivíduos, parece encontrar-se nos limites da zona ecológica submediterrânea (número 20 da Carta Ecológica [Albuquerque 1982]) nos casos a Norte de Évora, mas mais dentro da influência iberomediterrânea (zonas 21 e 22) nos restantes (figura 5.1).
Tendo em conta as características ecológicas (fito-edafoclimáticas) da localização dos povoamentos mistos estudados, pode considerar-se que estes são representativos, sob este critério, dos locais onde se encontraram híbridos (figura 5.1).
Conhecem-se numerosos pontos de contacto entre azinheira e sobreiro noutras regiões da Bacia Mediterrânica, como por exemplo no Alto Atlas [Belahbib et al. 2001], na Argélia e Sardenha [Sbay 2001] e por quase toda a Espanha suberícola [Jiménez & Gil 2001]. A inventariação de cerqueiros em todas as áreas de contacto entre as duas espécies que lhes dão origem poderia ter importância para definir factores ecológicos de risco da sua formação.
2) Presumíveis descendentes de híbridos
Houve poucos casos evidenciando a existência de linhagens derivadas dos híbridos, isto é, de indivíduos (sementes, plantas de viveiro ou adultos) cujos zimogramas sugerissem introgressão dum alelo de azinheira em genótipos de sobreiro (ou vice-versa).
A nível de adultos poderia citar-se o zimograma PGM “bd” na árvore 30 de Feijoas (azinheira), mas que, face à constatação de que a banda d de PGM pode ser o produto dum alelo raro presente em azinheiras, deve ser considerado normal (cf. “Resultados”, parte III, secção C1); de resto, registam-se o DIA “X” nas árvores 11 e 36 de Alfaiates (azinheiras) e alguns sobreiros de cortiça preguenta com DIA “X” ou “Az” em Montinho.
Em plantas jovens, depois de descontados os casos em que parece terem havido erros de avaliação de EST, contam-se apenas 2 azinheiras com PGM “d” (amostras 38c e 38i), algumas descendentes de A11 e A36 com o DIA “X” das respectivas mães, 2 sobreiros de Santiago do Cacém com DIA “X” (amostra 30AB) e “Az” (20K, que adicionalmente apresentava ACP-F intermédio entre sobreiro e azinheira), e 1 azinheira de Santiago do Cacém com zimogramas característicos duma ou outra das duas espécies, e também dos híbridos (amostra 15U).
As amostras 38c e 38i poderiam ser homozigóticos para o alelo de azinheira que codifica a banda PGM “d”, mas a improbabilidade desse genótipo (além de ser contraditório com o da presumível mãe, M19, expressando “ac”) é pouco aceitável no quadro da variação que se possa considerar normal em azinheira. É possível que de facto haja nestes dois casos um alelo de sobreiro introgredido, transmitido através do pólen, enquanto a actividade de azinheira (possivelmente a ou c, considerando o fenótipo da presumível mãe) não teria sido revelada.
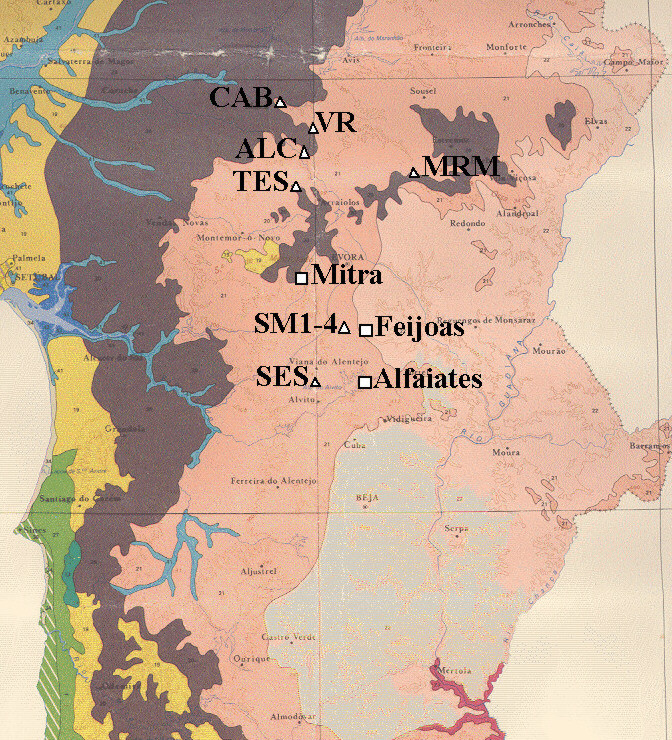 Figura 5.1 — Distribuição aproximada dos povoamentos
mistos (quadrados) e dos híbridos (triângulos) sobre a carta
ecológica [Albuquerque 1982]. As zonas ecológicas
submediterrânea (número 20 nesta carta), iberomediterrânea (22)
e mista (21) são, respectivamente, as de cor castanho-escuro, rosa
claro e rosa-carne.
Figura 5.1 — Distribuição aproximada dos povoamentos
mistos (quadrados) e dos híbridos (triângulos) sobre a carta
ecológica [Albuquerque 1982]. As zonas ecológicas
submediterrânea (número 20 nesta carta), iberomediterrânea (22)
e mista (21) são, respectivamente, as de cor castanho-escuro, rosa
claro e rosa-carne.Os casos de aparente introgressão em plântulas de sobreiro, tal como os dalguns sobreiros adultos de cortiça preguenta, são todos relativos ao zimograma DIA, que ou é “X” ou “Az”. O facto de coincidirem no locus de DIA pode sugerir o mesmo que para a banda d de PGM em azinheira, isto é, a possibilidade de existir em sobreiro um polipéptido de DIA variante, raro, que tem as mesmas propriedades na electroforese que o de azinheira; nesse caso, tratar-se-ia duma variação intraspecífica natural nesse locus, que ocorreria esporadicamente. Assim, a interpretação das ocorrências em Montinho é incerta: pode tratar-se dum local onde há indivíduos com o alelo supostamente de sobreiro, variante, e assim ser uma coincidência isso acontecer onde há sobreiros de cortiça preguenta; ou pode tratar-se duma verdadeira introgressão de genes de azinheira. Contudo, o facto de PER apresentar a banda 1 em proporção demasiado elevada para a espécie, nestes sobreiros e nos de Chaminé, já constitui um segundo indicador, aparentemente independente, de que esses sobreiros não são “puros”.
A observação de 2 zimogramas DIA “X” em sementes descendentes de A11, em contraste com a sua aparente ausência nas descendentes de híbridos, sugere que se trata de genótipos diferentes. Assim, coloca-se o mesmo problema que na interpretação dos zimogramas DIA “X” e “Az” em sobreiro: se se trata duma variante natural em azinheiras, ou da introgressão de genes de sobreiro.
Em suma, nenhum dos casos estudados, àparte talvez a amostra 15U, é um exemplo inequívoco de introgressão. Mas o contraste entre a sua frequência em árvores atípicas (sobreiros de cortiça preguenta) e a raridade nos povoamentos de referência e mistos, que se consideram representativos com base na sua composição e localização geográfica, favorece a hipótese de introgressão naquelas.
A raridade de exemplos de presumível introgressão nos povoamentos mistos e de referência pode ser explicada de duas formas, que não se excluem mutuamente:
i) o número insuficiente de marcadores de introgressão, pois teria de ser muito mais elevado (e distribuído uniformemente pelo genoma de sobreiro) para permitir uma sensibilidade adequada, o que parece estar fora do alcance de quem trabalhe apenas com marcadores isoenzimáticos — embora os mesmos não sejam de descartar em complementaridade com marcadores de DNA;
ii) tendência para as linhagens descendentes dos híbridos reaproximarem-se do padrão de cada
espécie, o que pode ou não envolver selecção disruptiva do património híbrido. A reduzida
proporção de descendentes de A11 e A36 com DIA “X” aponta nesse sentido, mas pode não
tratar-se dum alelo de sobreiro; na realidade, as descendências de SES, SM1 e SM2 apresentam
zimogramas de híbrido muito próximos dos esperados 50% por marcador, não dando por isso
qualquer evidência de selecção disruptiva. E como a maior parte dos descendentes dos híbridos
são retrocruzados, têm uma percentagem maioritária de genoma de sobreiro ou de azinheira,
o que provavelmente lhes confere uma preferência por continuarem a retrocruzar-se no mesmo
sentido (caso do SM2, cf. secção seguinte) e por isso as linhagens derivadas dos híbridos
tenderão a reaproximar-se dos genótipos característicos de sobreiro ou de azinheira
![]() .
.
3) Biologia da reprodução dos híbridos
Embora com alguma incerteza quanto à correcta identificação dos diversos zimogramas nos descendentes dos híbridos, o que poderá ter influenciado um pouco as distribuições na tabela 4.18, parece inequívoco que os cerqueiros têm capacidade de produzir descendência a partir de pólen de azinheira como de sobreiro, além duma percentagem apreciável de pólen recombinante, provavelmente de autopolinização. Assim, o surgimento de linhagens divergentes dos híbridos por retrocruzamento com as espécies que lhes deram origem pode processar-se nos dois sentidos opostos, isto é, aproximando de azinheira como de sobreiro.
As percentagens de pólen de sobreiro obtidas para SES, SM1 e SM2, a partir das plântulas deles descendentes da frutificação de 1998, foram bastante diferentes. No caso de SM2, poderá considerar-se que a alta preferência por pólen de sobreiro (83%) seja uma confirmação adicional de que se trata dum descendente de híbrido por retrocruzamento com sobreiro, isto é, o facto de já ter uma componente predominante de sobreiro, resultante desse presumível retrocruzamento, confere-lhe uma maior tendência para ser fertilizado por pólen desta espécie (mas sem excluir outros tipos de pólen). No caso de SES, nenhuma das poucas plântulas de 1998 terá derivado de pólen de azinheira, mas a análise em sementes do ano 2000 indica que não é incompatível com esta espécie. Apenas em SM1 houve em 1998 uma igual probabilidade de ser fertilizada por pólen de sobreiro e de azinheira.
Estes resultados são ainda preliminares. Uma caracterização mais completa, incluindo uma amostragem de mais híbridos e maior número de descendentes por cada árvore, e levando mais em conta algumas dificuldades de interpretação dos zimogramas destas descendências (por exemplo resolvendo a hereditariedade dos marcadores MDH+), é necessária para tentar estabelecer um padrão mais consistente do comportamento reprodutor dos híbridos.
Com a observação de que cerca de metade das descendentes de híbridos e pólen de azinheira recuperam caracteres das folhas da azinheira, pode ir-se ao ponto de tratar-se de apenas 1 locus para este fenótipo aparentemente "complexo", em que o alelo da azinheira é recessivo. A implicação é que, se se julgar uma azinheira apenas com base no aspecto das folhas, é possível que esse indivíduo possua 25% de genes de sobreiro na F2, ou até mais. Por extensão, outros caracteres em que o alelo recessivo seja o de sobreiro podem levar a considerar-se “sobreiros” indivíduos com uma elevada proporção de genes de azinheira (cf. figura 6.3, Apêndice II secção D). Se isso for verdade, então os eventos de hibridismo de há poucas gerações podem ser detectáveis com o pouco número de marcadores disponível, designadamente no que se trata de verificar nos sobreiros de cortiça preguenta ou os “negrais”. O que não exime da necessidade de caracterizar outros marcadores discriminantes.
1) Majorante da probabilidade de incidência de híbridos nos povoamentos mistos
Apesar do número bastante elevado de plantas provenientes dos povoamentos mistos analisadas, nenhuma apresentou as características dos híbridos, sendo todas concordantes com a espécie a que pertencia a árvore donde foram retiradas, aparte alguns casos de contaminação entre espécies que globalmente não chegaram a 1% (tabela 4.11). Este resultado negativo levaria a crer que não ocorreram híbridos nestas parcelas, mas é mais correcto majorar a probabilidade real com um valor P cujas estimativas se resumem na tabela 4.13.
Mesmo com o critério de risco menos rigoroso (α = 20%), o valor de P é inferior a 0,02% ou, se
se admitir que a formação de híbridos é principalmente através da fertilização das azinheiras com pólen
de sobreiro, este majorante pode subir até 0,043%. São valores suficientemente baixos para afirmar-se
que é uma ocorrência rara. Como nos sistemas de regeneração natural há que levar em conta todos os
processos de eliminação a que estas plantas (híbridas ou não) estão sujeitas, nomeadamente por acção
de herbívoros ou por desmoitas, e também o impacto da selecção natural (competição por recursos, por
exemplo), uma percentagem tão baixa implica que a ocorrência de um híbrido fértil nos povoamentos
deverá ser muito esporádica. Se adicionalmente considerar-se a possibilidade dalgumas desmoitas serem
selectivas contra os híbridos, reduz-se ainda mais a possibilidade de ocorrerem espontaneamente
![]() .
.
Isto entra em forte contradição com os estudos de polinização controlada que davam a entender uma elevada percentagem de híbridos [Boavida et al. 2001], mas cuja relevância, como se destacou anteriormente (“Revisão bibliográfica” parte II, secções C3 e D2), é relativa. E torna-se perfeitamente claro que outros resultados indicando taxas muito mais elevadas de hibridismo ou introgressão [Ellena-Rossellò et al. 1992, 1997, Toumi & Lumaret 1998] não são generalizáveis. Sendo um facto que os carvalhos cerqueiros resultam deste hibridismo (parte I), a sua ocorrência em Portugal contrasta com a grande extensão de contacto entre as duas espécies.
Como a totalidade das amostras de referência que serviram para definir o padrão “puro” de cada espécie eram plantas de viveiro, e os híbridos estudados tinham todos porte arbóreo, poderia colocar-se a hipótese da expressão dos marcadores de hibridismo ser diferente nas plântulas em relação aos adultos, e assim a aparente ausência de híbridos, na amostragem feita, mascarar uma ocorrência real mais elevada. Mas o aparecimento de padrões híbridos nas descendências dos cerqueiros (nomeadamente nas folhas de plantas jovens), assim como a total concordância da quase totalidade dos adultos dos povoamentos mistos com o padrão da respectiva espécie (à excepção de A11 e A36, cf. secção A), permite descartar tal hipótese — já em sementes, a ausência de zimogramas DIA “X” (e da banda 1 de PER), além do facto de GsR e PGM não poderem ser interpretados, constituem ainda uma forte limitação ao uso deste material no estudo do hibridismo.
Mas há que lembrar o facto da amostragem não ser muito representativa, e a vários níveis. Dentro de cada povoamento, apenas cerca de metade das descendências potenciais estavam representadas (tabela 3.1), o que em parte talvez fosse minorado se tivessem sido feitas várias colheitas de semente durante a época de frutificação, se se tivesse limitado o pastoreio nas parcelas, etc.; e houve famílias que o estiveram pouco, sendo relativamente fácil dentro de cada uma haver ocorrências de hibridismo não-detectadas (Apêndice IV secção A; mas cf. secção C1, para as azinheiras tardias); finalmente, há a possibilidade das árvores marcadas nos povoamentos mistos não serem susceptíveis de hibridar, em contraste com outras, destes ou doutros povoamentos. Em rigor, as estimativas de probabilidade aqui feitas aplicam-se apenas em materiais propagados em viveiro, nesse ano de frutificação (o contraponto com outros anos é analisado na secção C1) e, com limitações, nesses povoamentos. São, por isso, uma indicação inicial, mas que é suficientemente expressiva face aos pressupostos longamente reiterados de elevados riscos de hibridismo resultantes do contacto entre as duas espécies.
2) Implicações do valor obtido
Independentemente do valor real que alguma vez venha a ser obtido em cada povoamento misto, é do maior interesse examinarem-se as implicações da afirmação, qualitativa, de que este hibridismo é esporádico.
Aproveitamento para reprodução da semente de sobreiro proveniente de povoamentos mistos
A baixa probabilidade de formação de híbridos tornaria irrelevante a “boa prática” de rejeitar, para fins de propagação, as sementes de sobreiro provenientes de povoamentos mistos. Porém, na situação presente seria importante estudar mais extensivamente os materiais com essa proveniência, nomeadamente pela criação dum serviço de controlo da ocorrência de novos híbridos baseado nos marcadores desenvolvidos no presente trabalho (cf. secção D1). Esse mesmo serviço poderia até ser um meio de validar lotes de sementes para propagação provenientes de povoamentos mistos actualmente interditados pela actual legislação (decreto-lei 205/2003, anexo IX, parte A), o que num prazo relativamente curto se justificaria pela inexistência de argumentos objectivos para as restrições impostas e, ainda menos, para os limites fixados.
A adopção de medidas restritivas à utilização de semente para reprodução pode implicar enviesamentos muito perigosos do património genético de sobreiro que se vai propagando. Não só os povoamentos mistos abrangem uma área importante da distribuição do sobreiro em Portugal, como essa área corresponde a condições fito-edafoclimáticas que parecem ser especialmente propícias à produção da cortiça de melhor qualidade em sistema de montado. A prioridade a observar quanto à elegibilidade da semente de sobreiro de qualidade para reprodução, é a de haver um número suficiente de árvores reprodutoras por povoamento [Varela 1995], e que essa legislação não satisfaz apesar das intenções expressas no respectivo anexo III.
Em suma, deviam flexibilizar-se os critérios, incentivando os proprietários dos povoamentos mistos produtores de cortiça de boa qualidade, e com um número elevado de árvores, a candidatarem as sementes respectivas à qualificação como materiais de reprodução, na condição de os sujeitarem a um controlo sobre a ocorrência de híbridos.
Delineamento das amostragens para a detecção de hibridismo
Considerando à partida uma probabilidade muito baixa de ocorrência de híbridos, mesmo nas árvores que mais provavelmente estariam expostas ao pólen heterospecífico como era a hipótese inicial do presente trabalho, a etiquetagem família a família torna-se uma sobrecarga desnecessária. A primeira abordagem à detecção de híbridos formados em cada ano deve antes fazer-se por rastreios em larga escala, como por exemplo na realização de amostragens da produção dos viveiros ou de povoamentos onde se suspeite haver um maior risco de hibridismo (cf. secção D1); em complemento, por exemplo se se identificasse alguma proveniência com incidência apreciável de híbridos, ou árvores a priori mais propensas a hibridar por terem na sua proximidade apenas indivíduos da outra espécie, então justificavam-se estudos a nível familiar, necessariamente em larga escala, cujo interesse pode até transcender a temática do hibridismo (cf. secção C e parte III).
A ocorrência daquilo que os produtores do Alto Alentejo designam por “negrais” (a não confundir-se com a mesma designação para Quercus pyrenaica) não é de todo compatível com a estimativa feita. Segundo certos relatos, existem propriedades onde esta forma aparece na regeneração espontânea com bastante frequência (anualmente?) e em quantidade, propriedades essas que podem até não ter azinheiras na proximidade. Os negrais são sistematicamente desmoitados porque os produtores presumem tratar-se de híbridos. No decurso do presente trabalho não houve ocasião de analisar estas formas, mas é altamente desejável que essa análise se faça no futuro, com os marcadores genéticos disponíveis, de modo a tentar esclarecer o estatuto botânico destas plantas.
C — Factores que influenciam a probabilidade observada
Os valores de probabilidade de formação de híbridos são a resultante duma série de factores, limitadores ou facilitadores, que se passam a analisar no contexto do presente estudo e dos valores obtidos. Infelizmente, fora do projecto PAMAF 8153 que englobou o presente estudo, a informação relevante para esta análise é escassa, deixando muitas incógnitas por resolver.
Está largamente documentada a separação temporal entre as épocas de floração de sobreiro e azinheira, em que esta é a espécie mais precoce [Vasconcelos & Franco 1954, Varela 1995, Brandão 1996]. No entanto, a caracterização fenológica da floração em povoamentos marcados, como a que fez parte do projecto PAMAF 8153 (supervisionada pela Engª Maria Carolina Varela, da Estação Florestal Nacional), revela uma tal margem de variação intraspecífica, tanto em sobreiro como azinheira, que a simultaneidade das fases de polinização e receptividade de azinheiras tardias e sobreiros precoces é possível. Nomeadamente, observou-se no povoamento da Mitra, na Primavera de 1998 (de que resultou a frutificação do mesmo ano analisada no presente estudo), um período de pelo menos 14 dias em que 10 sobreiros (M01, M09, M14, M27, M32, M35, M40, M45, M47 e M50) produziam pólen em simultâneo com pelo menos duas de 5 azinheiras (M12, M48, M55, M60 e M64) [Brás 1999, Varela et al. em preparação].
O protandrismo destas duas espécies, embora não muito pronunciado [Vásquez 1998, Boavida et al. 2001], implica pelo menos em teoria que o período em que haveria oportunidade de polinização de azinheiras por sobreiro estender-se-ia para além dos referidos 14 dias — as 5 azinheiras mais tardias (e mesmo outras menos tardias) terão estado receptivas a pólen até doutros sobreiros menos precoces; esse mesmo protandrismo reduziria a possibilidade de polinização interspecífica no sentido inverso, mas durante esses 14 dias é provável que alguns dos sobreiros mais precoces também teriam estado receptivos quando ainda havia emissão de pólen de azinheira. Evidentemente, a situação torna-se ainda mais complicada tendo em conta que o pólen pode ter uma viabilidade prolongada (cf. “Revisão bibliográfica” parte II secção A2), e embora seja de admitir que o que está presente nas nuvens polínicas a cada momento é principalmente o libertado mais recentemente [Brandão 1996], a sua deposição na vizinhança dos estigmas antes destes estarem receptivos aumenta a probabilidade de polinização cruzada entre indivíduos com datas de floração menos próximas.
Os registos na Primavera de 2000 [Varela et al. em preparação], desta vez alargados às 3 parcelas mistas mas truncados (não houve possibilidade de acompanhar as florações depois de 5 de Abril, deste modo perdendo-se quase toda a informação sobre sobreiro), vieram revelar dois novos factos importantes: primeiro, que o comportamento de cada espécie era semelhante entre estes povoamentos, em termos de datas dos períodos de floração; segundo, que as datas em sobreiro foram de tal maneira adiadas, em relação às de azinheira, que a possibilidade de polinização interspecífica, mesmo de azinheira com pólen de sobreiro, pareceria quase nula: quase todas as azinheiras tinham terminado a sua fase de emissão de pólen (Fm2) em 5 de Abril, aproximadamente como em 1998, mas nessa altura apenas um sobreiro de Alfaiates atingira a fase E, e a maior parte nas 3 parcelas ainda não tinha floração para além da fase C, quando em 1998 os mais precoces da Mitra já se encontravam em Fm2 nessa data e, por isso, emitindo pólen quando havia azinheiras receptivas.
O paralelismo entre povoamentos em 2000 permite assumir-se que as datas de floração de 1998 em Feijoas e Alfaiates tiveram a mesma margem de variação que a registada na Mitra nesse ano; a ser assim, a frutificação de 1998 (representada nas plântulas caracterizadas para marcadores enzimáticos) teria sido das de maior oportunidade para o aparecimento de híbridos. Acrescente-se que o mesmo raciocínio se pode aplicar às nuvens polínicas de origem remota, uma componente importante da reprodução em Quercus: teria havido em 1998 um período onde conteriam uma elevada proporção de pólen heterospecífico.
Se considerarmos apenas as árvores cuja floração se deu durante os 14 dias de coexistência de pólen das duas espécies em 1998, obtiveram-se 137 descendentes de 4 azinheiras e 2 sobreiros, o que dá um valor de P (majorante da probabilidade de ocorrência de híbridos nas descendências, cf. secção B) de 0,2% com α = 20%, ou de 0,3% para as 89 descendentes de azinheiras. Mas deve notar-se que a maior parte dos sobreiros elegíveis para esta análise não produziram semente nesse ano — ou tê-lo-iam feito em data diferente da da colheita. Entretanto, se incluirmos as azinheiras de Feijoas e Alfaiates que correspondessem à classificação de tardias em 1998, a amostragem real deste grupo terá sido maior. Em 2000, as únicas azinheiras do povoamento da Mitra que em 5 de Abril ainda não tinham atingido a fase de ântese eram precisamente as mesmas 5 consideradas tardias em 1998, uma evidência de repetibilidade que já era conhecida em sobreiro [M. C. Varela, comunicação pessoal]; as azinheiras números 4 de Feijoas e 17 e 22 de Alfaiates foram igualmente tardias em 2000, e assumindo que também o teriam sido em 1998 então podem acrescentar-se as 10 descendentes de F04 (não se obtiveram descendentes de A17 ou de A22), o que reduz P para 0,2% com α = 20%.
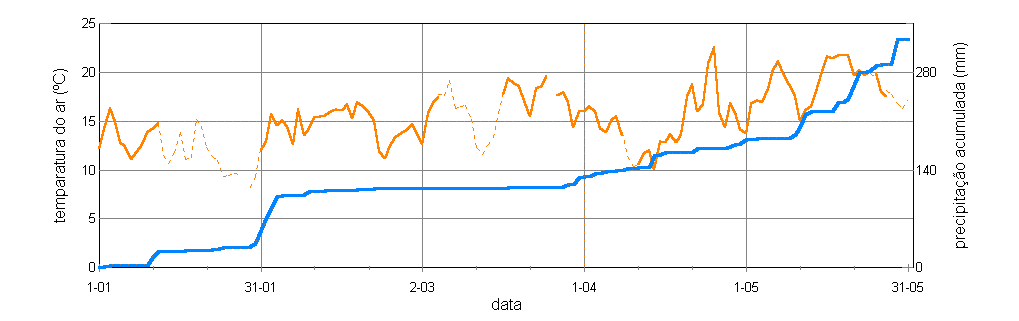
As variações nalguns parâmetros climáticos, de ano para ano, poderiam ser uma causa para as diferenças observadas entre estes dois anos. Os registos do Centro Geofísico de Évora (http://www.cge.uevora.pt) permitem analisar estes parâmetros (figura 5.2). Das diferenças observáveis, parecem de especial relevância, em 1998, as temperaturas do ar relativamente altas nos dois primeiros meses, e em 2000 a reduzida precipitação até ao final do Inverno (acumulação de apenas 23 mm até 19 de Março). É plausível que essas temperaturas de 1998, acompanhadas duma acumulação precoce de precipitação na ordem dos 110 mm, tivessem facilitado uma antecipação da floração de sobreiro nesse ano — ou a falta de água até final do Inverno de 2000 (e as temperaturas frias de Janeiro) a tivessem retardado. A comprovar-se uma dependência das datas dos períodos de fertilidade de factores climáticos, que no caprichoso clima do Alentejo poderia manifestar-se tanto nas azinheiras [Brandão 1996] como nos sobreiros [Brás 1999, M. C. Varela et al. em preparação], então é previsível que no decurso da longa vida fértil de cada árvore a oportunidade de polinização interspecífica varie muito de ano para ano.
a)
b)
 Figura 5.2 — Registos da média diária da temperatura do ar (linha laranja) e da precipitação
acumulada (linha azul) relevantes para o povoamento da Mitra. a) Ano de 1998. Os registos na estação
meteorológica da Mitra são incompletos, tendo as lacunas sido extrapoladas a partir dos valores de
Reguengos, no caso das temperaturas somando a média das diferenças diárias (2,7 ºC) entre as duas
estações nesse ano (troços a tracejado), e no caso da precipitação usando directamente os valores de
Reguengos, que noutros anos foram muito semelhantes aos da Mitra. b) Ano de 2000.
Figura 5.2 — Registos da média diária da temperatura do ar (linha laranja) e da precipitação
acumulada (linha azul) relevantes para o povoamento da Mitra. a) Ano de 1998. Os registos na estação
meteorológica da Mitra são incompletos, tendo as lacunas sido extrapoladas a partir dos valores de
Reguengos, no caso das temperaturas somando a média das diferenças diárias (2,7 ºC) entre as duas
estações nesse ano (troços a tracejado), e no caso da precipitação usando directamente os valores de
Reguengos, que noutros anos foram muito semelhantes aos da Mitra. b) Ano de 2000.
2) Barreiras reprodutoras após a polinização
Uma vez aceite que existe a possibilidade dum estigma receptivo de azinheira receber grãos de pólen viáveis de sobreiro (e não excluindo a combinação recíproca), até que ponto irá ou não resultar em fertilização, e depois no desenvolvimento e maturação da semente? Sabendo que a maior parte dos gametófitos masculinos é eliminada antes da conjugação, esta questão pode reformular-se da seguinte maneira: até que ponto essa eliminação é diferencial entre os da própria espécie e os da outra espécie?
No limite, poderia considerar-se que a probabilidade média dum grão de pólen da outra espécie desenvolver um tubo polínico, e a deste chegar aos ovários e fertilizar é a mesma que a dum grão de pólen da mesma espécie; isso significaria não haver, na fase pré-zigótica, barreiras reprodutoras específicas à hibridação. Entretanto, o presente estudo foi feito durante um ano de frutificação que, tendo em conta os estudos de floração efectuados em paralelo (cf. secção anterior), faria esperar uma elevada taxa de hibridação em algumas descendências pelo menos, caso não houvesse barreiras reprodutoras específicas à hibridação.
Mas se de facto o grão de pólen da outra espécie tiver em média menos probabilidade de germinar no estigma receptivo do que os da própria espécie, e/ou se o subsequente tubo polínico for em média mais facilmente travado no seu desenvolvimento antes de chegar ao ovário, então há que acrescentar este factor de redução da probabilidade de hibridação, e a ausência de híbridos nas plântulas obtidas dos povoamentos mistos em 1998 parece corroborá-lo. Embora os estudos experimentais com polinização controlada tendam a apoiar esta possibilidade [Boavida et al. 2001], a realidade é que ainda não foi abordada directamente, pela inclusão duma mistura de pólen das duas espécies. Isso não permite, nomeadamente, obter uma estimativa quantitativa da possível redução do sucesso da polinização interspecífica durante a fase pré-zigótica, que constituiria uma referência importante para o estudo de factores de risco de hibridação.
Mas, para além da probabilidade média há que contar com possíveis variações consoante os indivíduos envolvidos: da mesma maneira que há diferentes graus de compatibilidade entre indivíduos da mesma espécie [Yacine & Bouras 1997], também essa variação pode existir entre os de espécies diferentes [Kanazashi et al. 1997b]. Por isso é de admitir que a ocorrência de híbridos seja mais previsível quando se encontram próximos entre si um sobreiro e uma azinheira compatíveis e cuja floração, pelo menos nalguns anos, se sobreponha no tempo. Esta hipótese, sem excluir a possibilidade de ocorrerem híbridos por polinização a longas distâncias, poderia fornecer uma pista para a ocorrência de híbridos em apenas algumas áreas com povoamentos mistos (figura 5.1).
Deste modo, um dos critérios de elegibilidade de povoamentos de sobreiro para aproveitamento de semente para propagação, que é o de haver quando muito 15% de outras quercíneas (decreto-lei 205/2003, anexo IX, parte A), além de ser evidentemente arbitrário, pode revelar-se inútil: mesmo numa percentagem abaixo dessa, e dependendo das condições particulares que facilitem a polinização interespecífica, poderia ocorrer hibridismo.
Há ainda a possibilidade daquilo que ficou conhecido como “emasculação ambiental” em que, sob certas condições ambientais, a viabilidade do pólen da própria espécie ser de tal modo afectada que as flores femininas acabam por ser fertilizadas a partir de pólen doutra espécie. No contexto dos povoamentos mistos de sobreiro e azinheira em Portugal, tal fenómeno parece não ocorrer, até porque resultaria em povoamentos onde predominam híbridos, como no exemplo original [Williams et al. 2001], o que não parece acontecer com os cerqueiros em ponto algum da Bacia Mediterrânica.
Após a fertilização, existe a possibilidade de eliminação preferencial dos híbridos durante o desenvolvimento do embrião, durante o período após a deiscência até à germinação, durante a germinação ou durante o desenvolvimento da plântula. Não parece existir qualquer informação disponível, nem mesmo extrapolada doutros híbridos de Quercus, a este respeito. Mas é provável que haja a eliminação sistemática dos cerqueiros jovens nas desmoitas (se se trata dos “negrais”, cf. secção B, terá de ser averiguado) em povoamentos onde só se pretende a regeneração de sobreiro. E existe a possibilidade da ocorrência dos híbridos coincidir com a ausência destas desmoitas selectivas.
D — Ideias para projectos de futuro
Dispondo-se de marcadores isoenzimáticos dos híbridos entre sobreiro e azinheira, abre-se a possibilidade de lançar linhas de investigação sobre várias questões relacionadas. Nesta secção revêem-se algumas dessas possibilidades, dando ênfase à maneira como se articulam com os resultados do presente trabalho e ao papel que a análise destes marcadores pode desempenhar.
1) Incidência de hibridismo em materiais de propagação
O potencial dos marcadores isoenzimáticos para análises em larga escala tem a sua aplicação mais evidente na amostragem dos materiais de propagação de sobreiro de proveniências portuguesas, especialmente aquelas que abrangem povoamentos mistos com azinheira. Os povoamentos mistos ocorrem um pouco por toda a distribuição de sobreiro em Portugal (tabela 5.1), e grande parte das proveniências contribuem para o material de propagação comercializado por viveiristas portugueses.
Tabela 5.1 — Distribuição dos povoamentos mistos em Portugal, por unidades territoriais, segundo uma estimativa feita em colaboração com a Associação Nacional de Empresas Florestais Agrícolas e do Ambiente (ANEFA), em 2001. As regiões de proveniência de sobreiro são as definidas por Varela [1997].
Região agrária |
Região de proveniência |
Unidades territoriais |
Trás-os-Montes |
III |
Douro Superior (1%), Nordeste (9%) |
Beira Litoral |
II |
Pinhal litoral (2,5%), Pinhal e Beira Serra (2,5%) |
Beira Interior |
V |
Pinhal Interior Sul (5%) e Beira Interior Sul (5%) |
Ribatejo e Oeste |
IV |
Tomar (4,5%), Abrantes (4,5%), Chamusca e Coruche (6%) |
Alentejo |
IV |
Alentejo litoral (5,5%) |
Alentejo |
V |
Alto Alentejo (16,5%), Alentejo central (22%), Baixo Alentejo (11%) |
Algarve |
VII |
Barlavento (1,5%), Sotavento (3,5%) |
Essa amostragem poderia ser feita ao longo do ano já com as plantas em pleno desenvolvimento. Visar-se-ia, numa primeira linha, a identificação de híbridos com um conjunto de marcadores diagnosticantes suficiente — a escolha recaindo provavelmente sobre DIA e MDH, no sistema de separação C, que deram provas de elevada consistência e facilidade de interpretação dos fenótipos “X”. Não haveria grande constrangimento logístico, com o necessário investimento em recursos humanos, para processar em poucos meses alguns milhares de amostras de cada ano (segundo os cálculos do projecto PAMAF 8153, realiza a extracção e análise no sistema C, 450 amostras por mês para um técnico). A incidência sobre povoamentos mistos seria particularmente relevante no esclarecimento dos pressupostos para a certificação de semente como seleccionada (como definido no anexo IX, parte A, do decreto-lei 205/2003, cf. secções B e C2 e “Introdução”), entretanto em vigor, que é obrigatória para os materiais de propagação de sobreiro (artigo 17º desse decreto). Mas não é de excluir a utilidade de estender o rastreio a materiais de povoamentos “puros”, incluindo de proveniências estrangeiras que sejam criadas em viveiros portugueses.
Uma vez encontradas plantas com zimogramas de híbridos, outros marcadores têm de ser investigados para verificar esse hibridismo em todos, o mesmo tendo de fazer-se para as restantes amostras das respectivas proveniências, o que no espaço de 1 mês (ou até mais), usando a metodologia desenvolvida no presente trabalho, não requer a produção de mais extractos dos mesmos indivíduos.
A partir daqui, pode considerar-se prioritário, ou não, intensificar a amostragem de materiais de viveiro das proveniências em causa, ou fazê-lo apenas para as frutificações dos anos seguintes. A análise dos registos climáticos mais próximos geograficamente, na Primavera anterior, é importante para servir de padrão para futuras observações da fenologia da floração nas duas espécies em cada proveniência, acompanhadas por registos edafoclimáticos durante cada época, para depois recolher-se semente (nos sobreiros e nas azinheiras) na respectiva época de frutificação. Como potencial factor de risco de hibridação (“emasculação ambiental” [Williams et al. 2001]), a viabilidade do pólen de azinheira (e eventualmente de sobreiro) deve ser investigada nestes povoamentos. A viabilidade e sobrevivência em viveiro das descendências recolhidas destes povoamentos também deve ser registada.
Os resultados principais a esperar dum tal estudo são a identificação de proveniências onde ocorrem plantas híbridas que podem ser vendidas em viveiro, e a estimativa de quantas ocorrem nos diversos anos. Complementarmente, há a oportunidade de caracterizar morfologicamente esses híbridos em fase de plântula, caso seja divergente da descrição botânica [Vasconcelos & Franco 1954], e a de aumentar o conhecimento sobre as condições edafoclimáticas que facilitam a polinização interspecífica.
Além disso, pelo facto de também se estudarem as condições locais de formação de híbridos, esta linha de trabalho permite uma melhor definição dos factores de risco de hibridação entre sobreiro e azinheira, que pode ter reflexos positivos na gestão dos povoamentos mistos e do aproveitamento da semente neles produzida.
2) Biologia da reprodução dos cerqueiros
Quanto maior for o número de carvalhos cerqueiros inventariados, mais relevantes serão as conclusões que se podem tirar sobre a sua fertilidade com pólen de sobreiro e de azinheira, que para já só conta com dados bastante limitados, havendo até a possibilidade duma das árvores (SM2) não ser um híbrido mas sim descendente de híbrido. Seria desejável obter em cada ano algumas dezenas de plantas descendentes de cada híbrido (o que pode não ser fácil nalgumas árvores) e analisar o máximo número possível de marcadores enzimáticos, complementados por outros marcadores genéticos (cf. secção 5). As questões relacionadas com a viabilidade do pólen dos cerqueiros e a germinação e sobrevivência em viveiro das suas descendências podem ser relevantes.
Também faz parte da biologia da reprodução dum híbrido estudar as descendências de azinheiras ou sobreiros na sua imediata vizinhança, sobretudo se se presumir que a receptividade das flores femininas dalgumas dessas árvores coincide com a emissão de pólen recombinante pelo híbrido — é de presumir que uma parte relevante do pólen que essas árvores recebem provém do híbrido que lhe está próximo. Esta abordagem justifica-se também para compensar a falta de informação disponível nos híbridos em que a produção de semente seja reduzida.
Os resultados da análise isoenzimática das descendências dos híbridos permitem testar se em todos os híbridos existe uma distribuição igual entre pólen de sobreiro e de azinheira, estimar a percentagem de pólen recombinante que recebem, verificar se o aparecimento de descendentes de pólen dos híbridos nas árvores vizinhas depende da coincidência entre datas de floração, e ainda testar a ligação cromossómica entre os loci discriminantes (enzimáticos e outros).
Se em paralelo for realizada a linha de trabalho descrita na secção anterior, pode vir a dispor-se duma grande riqueza de informações que conduzam ao melhor conhecimento da dinâmica de formação de híbridos e subsequente manutenção de introgredidos nos povoamentos (cf. parte I secção A).
Finalmente, pode perspectivar-se transferir estas plantas para um local definitivo, para acompanhar o desenvolvimento da cortiça e tentar correlacionar variações na sua expressão com a presença de certos marcadores genéticos (isoenzimáticos ou outros) ou de certas variantes alélicas de genes candidatos e sua expressão no felogene. Nomeadamente, tentar-se correlacionar com a cortiça preguenta de modo a determinar o grau de introgressão, se de facto é o que se trata, envolvido na sua formação (cf. secção seguinte). Evidentemente, por precaução, estes “pomares de notomorfos” teriam de encontrar-se a uma distância suficientemente grande (talvez 10 km [Streiff et al. 1999]) de sobreiros ou azinheiras, para evitar cruzamentos indesejáveis.
3) Investigação dos sobreiros de cortiça preguenta
O presente estudo apenas aflorou a análise dos sobreiros de cortiça preguenta, como possível aplicação dos marcadores discriminantes. Não havendo nada de conclusivo, há porém dados suficientes para delinear essa aplicação de maneira mais incisiva.
Na hipótese destes sobreiros terem introgressão de genes de azinheira, a sensibilidade da detecção depende do número de marcadores genéticos diagnosticantes, que idealmente terão de ser muitos e distribuídos uniformemente pelo genoma. O facto de ter-se detectado o que parece ser introgressão, no locus de DIA e possivelmente num dos loci de PER, sendo tão poucos os marcadores utilizados neste estudo, se não for um golpe de sorte é porque a percentagem do genoma de sobreiro não precisa de ser muito próxima de 100% para um sobreiro parecer como tal; nesse caso, e tendo em conta que o extenso contacto que existe actualmente entre as duas espécies não deve ser anterior às últimas décadas do século XIX (“Revisão bibliográfica”, parte III, secção A), a separação entre eventos de hibridismo e os actuais descendentes de híbridos que possam existir não deve ir além de 2 gerações. A ser verdade, os marcadores enzimáticos podem ter a sensibilidade necessária, mas evidentemente têm de ser complementados por outros, e é de grande importância incluir caracteres morfológicos, para além da qualidade da cortiça, que em paralelo devem assinalar introgressão caso a mesma exista (cf. tabela 2.3, “Revisão bibliográfica” parte III secção C3).
Como referido na secção anterior, a análise dos notomorfos nos locais definitivos pode vir a trazer pistas sobre quais os loci mais importantes para investigar nos sobreiros de cortiça preguenta.
Em termos de abordagem experimental, a investigação do estatuto genético dos “negrais” seria idêntica; em termos de amostragem, geralmente não sobrevivem para além duma fase precoce no campo, precisamente porque se distinguem morfologicamente dos sobreiros típicos. Se houver “negrais” adultos, então pode averiguar-se até que ponto eles se distinguem da norma nesta fase, nomeadamente em termos de qualidade da cortiça que produzem. Alternativamente, a diferença morfológica dos “negrais” pode ser uma manifestação precoce de plasticidade fenotípica, sem relacionar-se com a introgressão de genes de azinheira.
4) Inventariação de híbridos e seus descendentes
É importante continuar-se o inventário de carvalhos cerqueiros, não só pelo valor que tem em si mesmo mas pela possibilidade que dá de cada vez melhor definir os factores de risco de hibridação entre sobreiro e azinheira. A esse inventário também interessará associar um outro, decorrente das análises que se venham a realizar, de indivíduos, povoamentos e unidades territoriais onde incide introgressão de azinheira em sobreiro (e também vice-versa). Todos os indivíduos inventariados devem ser analisados para o máximo número possível de marcadores diagnosticantes.
5) Complementaridades a cultivar
Os marcadores de DNA podem complementar os enzimáticos, não só porque acrescentam outras potencialidades, mas porque provavelmente se localizam em pontos do genoma diferentes e assim podem preencher lacunas de sensibilidade destes últimos, nomeadamente para a análise de indivíduos introgredidos (plantas de viveiro, descendentes dos híbridos e das árvores que os circundam, sobreiros de cortiça preguenta). Os progenitores de cada híbrido — seja os que se encontram no campo, especialmente se forem relativamente jovens, seja os que se detectam nos materiais de viveiro — podem eventualmente ser encontrados na proximidade, para tal sendo fundamental a utilização de análises em muitos loci (designadamente, AFLPs) ou em loci com grandes séries alélicas (nSSRs [Soto et al. 2003] e SSCPs), e a diagnose do progenitor feminino (cpDNA).
O principal problema com estes marcadores é o custo por amostra e o tempo de realização, que podem ser proibitivos em contextos como o de rastreios em larga escala. Segundo a questão onde podem ser aplicados, e as respectivas exigências, assim a prioridade maior será para os marcadores enzimáticos ou dos marcadores de DNA. Em qualquer das situações que se coloque, não pode menosprezar-se a complementaridade entre uns e outros.
Nas secções que precedem, os estudos fenológicos da floração em sobreiro e azinheira são constantemente referidos, pois como se demonstrou na análise dos resultados do projecto PAMAF 8153, constituiram importante complemento para a sua interpretação (cf. secção C). É um exemplo paradigmático da sinergia que resulta de duas abordagens ao mesmo problema, neste caso a reprodução sexuada em sobreiro e azinheira em povoamentos mistos. Porém, deve realçar-se a intensidade com que os recursos humanos se têm de envolver durante a época de floração para obter resultados tão detalhados como os do povoamento da Mitra em 1998.
Outra complementaridade vem dos estudos com polinizações controladas. Esta abordagem é a que permite compreender os factores envolvidos na eliminação dos tubos polínicos (cf. “Revisão bibliográfica” parte II, secção C). São muitas as abordagens já exemplificadas na bibliografia existente, todas com méritos importantes mas nenhuma delas considerando um aspecto fundamental: a aplicação de pólen de sobreiro e azinheira misturados. Note-se que até ao presente estudo não se dispunha de marcadores genéticos que permitissem identificar a origem do grão de pólen para cada descendente, mas existindo agora essa possibilidade, com marcadores enzimáticos pelo menos, a utilização de misturas de pólen é incontornável. Também devem resolver-se as dificuldades experimentais encontradas na polinização de sobreiro [Boavida et al. 2001]. E deve ter-se em conta a verificação de variações na compatibilidade entre indivíduos das duas espécies, na presunção de ser um factor de risco de hibridação (secção C2).
6) Precauções importantes a priori
O projecto PAMAF 8153 definiu um protocolo de trabalho que permitiu responder a algumas questões com bastante clareza, e que devem ser levadas em conta no planeamento doutros estudos: selecção de povoamentos representativos, complementaridade com estudos da floração no campo, criação das descendências em viveiro, extracção dos enzimas das folhas.
Por outro lado, houve aspectos da amostragem que não puderam ser corrigidos, e que requerem a maior atenção caso se pretenda uma análise familiar: a colheita de sementes já caídas no solo deve ser abolida (no caso das copas muito altas, pode cobrir-se o chão com um pano e varejar a folhagem), cada povoamento deve ser visitado várias vezes em cada época de frutificação para que não se excluam famílias inteiras, e o número de descendentes por árvore deve ser uniformizado.