Parte III: Híbridos de sobreiro com azinheira em Portugal e na Bacia Mediterrânica
A — Perspectiva histórica do problema
A formação de híbridos entre duas espécies requer uma sobreposição geográfica, entre populações
dessas espécies, mais ou menos extensa (por outras palavras, as espécies devem ser parapátricas ou
simpátricas). No caso do sobreiro e da azinheira (doravante este termo refere-se à subespécie
rotundifolia, excepto onde se expresse o contrário), a ocorrência de ambas em Portugal, Espanha, França,
Marrocos e Argélia permite em princípio esse contacto, como aliás o confirma Natividade [Subericultura
![]() pp. 37–73] e vem documentado mais recentemente [Toumi & Lumaret 1998]
pp. 37–73] e vem documentado mais recentemente [Toumi & Lumaret 1998]
![]() . Porém, as exigências
ecológicas de cada uma são bastante diferentes, levando a que esse contacto, pelo menos em condições
naturais, deva ser muito limitado: assim, em comparação com a azinheira, o sobreiro é menos tolerante
à secura e ao frio intenso, além de ser calcífugo [Bellarosa 2003], enquanto a azinheira evita locais com
maior humidade, para o que deve contribuir, pelo menos em parte, uma maior susceptibilidade ao
parasitismo em condições de alagamento [Sánchez et al. 2005]. Talvez se possa considerar que as
distribuições quase disjuntas registadas no Magrebe [Bernard 1937], pela presumível ausência de
políticas de povoamento sistemático por qualquer uma das espécies, sejam um testemunho da presumível
tendência destas duas espécies para não ocorrerem nos mesmos locais.
. Porém, as exigências
ecológicas de cada uma são bastante diferentes, levando a que esse contacto, pelo menos em condições
naturais, deva ser muito limitado: assim, em comparação com a azinheira, o sobreiro é menos tolerante
à secura e ao frio intenso, além de ser calcífugo [Bellarosa 2003], enquanto a azinheira evita locais com
maior humidade, para o que deve contribuir, pelo menos em parte, uma maior susceptibilidade ao
parasitismo em condições de alagamento [Sánchez et al. 2005]. Talvez se possa considerar que as
distribuições quase disjuntas registadas no Magrebe [Bernard 1937], pela presumível ausência de
políticas de povoamento sistemático por qualquer uma das espécies, sejam um testemunho da presumível
tendência destas duas espécies para não ocorrerem nos mesmos locais.
Em Portugal o sobreiro e a azinheira coexistem numa extensa faixa de sobreposição grosso modo equivalente à transição entre as regiões submediterrânica e iberomediterrânica (número 21 na legenda da Carta Ecológica de Portugal Continental [Albuquerque 1982]), que abrange áreas importantes desde a Beira Baixa até ao Algarve, para além doutras zonas mais limitadas, nomeadamente em Trás-os-Montes e Alto Douro. Esta vasta sobreposição é facilitada pela intersecção entre os nichos climáticos de ambas as espécies [M'Hirit 1999] e terá sido mais ou menos importante consoante as tendências históricas [Subericultura pp. 39–49], mas a sua actual vastidão parece resultar sobretudo de evoluções bastante recentes, isto é a partir do final do século XIX, no que respeita ao aproveitamento dos terrenos na metade Sul do território continental.
Até ao século XVIII, isto é, antes da generalização do uso do vidro no engarrafamento das bebidas,
o interesse do sobreiro residia essencialmente no aproveitamento do lenho (construção naval e carvão),
não-renovável, e ainda o do entrecasco (curtimenta) e da cortiça virgem (flutuadores), de valor
secundário em relação à exploração da bolota doce, entre outros usos, da azinheira [Natividade 1947,
Mendes 2002, Coelho 2003]
![]() . A situação de recuo do sobreiro na bacia hidrográfica do Tejo, motivado
pela procura de lenha para carvão [Natividade, 1947] assim como pela construção civil que se seguiu ao
terramoto de 1755 [Mendes 2002], teria provavelmente retirado o sobreiro de qualquer contacto que
tivesse tido com os montados de azinho no início do século XIX. Mas a escassez da cortiça amadia para
a indústria vinícola nas primeiras regiões de produção de rolha, sobretudo a Catalunha, deu origem a uma
procura deste recurso renovável, que foi aumentando ao longo do século XIX, com um melhoramento
progressivo da gestão dos povoamentos [Subericultura p. 46], embora parcimoniosamente [Mendes
2002], só havendo a certeza duma expansão da sua área geográfica a partir do terço final do século XIX,
passando-se de 121000 ha em 1867 para 651400 ha em 1950 [Mendes 2002, Coelho 2003], no que teve
paralelo na azinheira até à chamada campanha do trigo (os terrenos incultos no Alentejo desapareceram
praticamente no final do século XIX), sempre na sombra do interesse na produção cerealífera e em
estreita dependência da mão-de-obra seareira, que também era aproveitada para a extracção da cortiça
[Mendes 2002]. O consequente aumento do “capital suberícola” português distingue-se completamente
da expansão do pinheiro bravo (Pinus pinaster Ait.) nas três primeiras décadas do século XX,
comparativamente repentina [Mendes 2002], e expressa-se no aumento de produção e exportação de
cortiça a partir das décadas à volta de 1900 [Subericultura p. 49, Raposo s.d.
. A situação de recuo do sobreiro na bacia hidrográfica do Tejo, motivado
pela procura de lenha para carvão [Natividade, 1947] assim como pela construção civil que se seguiu ao
terramoto de 1755 [Mendes 2002], teria provavelmente retirado o sobreiro de qualquer contacto que
tivesse tido com os montados de azinho no início do século XIX. Mas a escassez da cortiça amadia para
a indústria vinícola nas primeiras regiões de produção de rolha, sobretudo a Catalunha, deu origem a uma
procura deste recurso renovável, que foi aumentando ao longo do século XIX, com um melhoramento
progressivo da gestão dos povoamentos [Subericultura p. 46], embora parcimoniosamente [Mendes
2002], só havendo a certeza duma expansão da sua área geográfica a partir do terço final do século XIX,
passando-se de 121000 ha em 1867 para 651400 ha em 1950 [Mendes 2002, Coelho 2003], no que teve
paralelo na azinheira até à chamada campanha do trigo (os terrenos incultos no Alentejo desapareceram
praticamente no final do século XIX), sempre na sombra do interesse na produção cerealífera e em
estreita dependência da mão-de-obra seareira, que também era aproveitada para a extracção da cortiça
[Mendes 2002]. O consequente aumento do “capital suberícola” português distingue-se completamente
da expansão do pinheiro bravo (Pinus pinaster Ait.) nas três primeiras décadas do século XX,
comparativamente repentina [Mendes 2002], e expressa-se no aumento de produção e exportação de
cortiça a partir das décadas à volta de 1900 [Subericultura p. 49, Raposo s.d.
![]() p. 97], assim como na sua
progressiva estabilização a partir dos anos 40 do século XX [Raposo s.d.4 apêndice F (2º)], com um
máximo de extracção de cortiça virgem a registar-se nos anos 60 [Mendes 2002].
p. 97], assim como na sua
progressiva estabilização a partir dos anos 40 do século XX [Raposo s.d.4 apêndice F (2º)], com um
máximo de extracção de cortiça virgem a registar-se nos anos 60 [Mendes 2002].
Segundo Raposo [s.d.4 pp. 94–95], este desenvolvimento veio na sequência da reabilitação dos solos com a aplicação sistemática de superfosfato de Cálcio, orientada para a produção de cereal mas que, com o declínio desta produção que se seguiu, deu a indicação, através da vigorosa regeneração espontânea de sobreiro que se observava, do porquê da anterior incapacidade desses solos para sustentarem arvoredo, o que acabou por ser uma realização da “profecia” de Barros Gomes em 1875, duma recuperação da antiga dominância do sobro (e azinho) no Alentejo, à altura restrito a pequenos povoamentos [Neves 1944, Raposo s.d.4 p. 96]. Morais considerava que a distribuição do sobreiro que se observava diferia substancialmente daquela onde naturalmente predominaria, pela cedência do seu terreno de anterior domínio, próximo do litoral a Norte do Tejo, para o pinheiro-bravo, mas tornando-se abundante em zonas de azinheira [Morais 1940]. Como complemento desta ideia, Amorim Girão [1942] apresentou um mapa dos topónimos portugueses relacionados com o sobreiro, e cuja concentração em Entre Douro e Minho, nas bacias do Mondego e do Zêzere, e na Estremadura, sugere uma distribuição em tempos antigos muito diferente da actual, resultando essa diferença do depauperamento das áreas florestais nessas zonas, para favorecer a agricultura e o povoamento [Subericultura pp. 43–44], por um lado, e pela expansão no Sul que, como acaba de apontar-se, seria relativamente recente.
Entretanto, a expansão do sobreiro nas regiões interiores de Portugal, para a produção de cortiça amadia, não desalojou a azinheira porque esta última reteve, até bastante tarde no século XX, o interesse económico que já tinha, ao qual se acrescentou cada vez mais a produção de carvão [Nogueira 1978, Coelho 2003], factor ao qual se pode ainda associar a garantia de salvaguarda económica que conferia às explorações a melhor resistência da azinheira aos rigores climáticos das regiões interiores. Assim, a extensão actual dos montados mistos terá sido a resultante dum compromisso entre a busca de rendimentos dum novo produto (a cortiça amadia) e a manutenção das fontes de rendimento anteriormente mais relevantes.
Embora as ligue uma relação causa-efeito, também é inegável a desproporção entre a actual extensão de montados mistos de sobreiro e azinheira em Portugal e a ocorrência espontânea dos híbridos entre estas duas espécies: face às oportunidades de hibridação nesses montados, esta deve considerar-se rara, mesmo descontando a possível contribuição de eventuais desbastes selectivos para estas árvores. Tendo em vista a marcada contracção da área com sobreiro e a predominância de terrenos incultos até ao terço final do século XIX, a sobrevivência de linhagens derivadas de híbridos anteriores à exploração intensiva da cortiça amadia deve ter sido excepcional. Ou seja, a considerar-se a introgressão de genes de azinheira nos sobreiros actuais, ela não é particularmente antiga: à escala do ciclo reprodutor do sobreiro (mínimo de 15–20 anos para frutificar, com muita irregularidade da produção de semente [Subericultura p. 103]), as linhagens híbridas formadas a partir da expansão do sobreiro, a existirem, remontam a menos de 8 gerações, e provavelmente o real número tenderia a ser muito inferior. Daí que a introgressão de azinheira em sobreiro, onde possa existir, tenderá assim a ser ainda elevada (depois de uma geração de retrocruzamento com sobreiro é em média, teoricamente, 25%, após duas gerações 12,5%, etc., mas ver Apêndice II secção D). Estes pressupostos contrastam com a linha de raciocínio de Natividade, quando justificava o polimorfismo nos sobreiros na variedade de combinações genotípicas que podem resultar das linhagens destes híbridos, após “inumeráveis” gerações [Natividade 1934 p. 126].
B — Implicações silvícolas e “peso” no sector
1) A produção de cortiça e o problema da qualidade
Estima-se que actualmente o sector da cortiça movimente em Portugal cerca de 900 milhões de Euros anuais, com uma taxa de cobertura exportação/importação de 640% [DGF 2003]. Com base no Inventário Florestal, estima-se que a produção em Portugal ronde as 105 mil toneladas anuais, cerca de 50% do total mundial, quota que atinge os 80% no que respeita à transformação, da qual 95% é exportada [DGF 2003]. Apesar deste excedente de procura por parte da indústria transformadora, a evolução dos custos de exploração e a necessidade de planear com décadas de avanço leva os diversos interessados no sector primário a preocuparem-se com a sustentabilidade desta exploração. As razões são várias, muito em função das perspectivas de cada um:
• Substituição da cortiça para a indústria vinícola por materiais alternativos (nomeadamente rolhas de plástico ou de alumínio) e consequente redução da procura;
• Declínio de produção da cortiça amadia por envelhecimento dos montados e falta de regeneração;
• Prejuízos associados à exploração de sobreiro, provocados seja por doenças e pragas, seja por introdução de germoplasma criado em viveiro sem controlo das proveniências, com os consequentes problemas de adaptação;
• Substituição do sobreiro, nas áreas onde actualmente se encontra distribuído, por outros tipos de exploração.
A complexidade destas preocupações torna o futuro da exploração da cortiça em Portugal problemático. Mas se há factor que é relevante para todas elas, é a qualidade da cortiça produzida. Embora se reconheça não existirem ainda materiais alternativos para o rolhamento de bebidas engarrafadas que a igualem sequer, isso não quer dizer que a superioridade da cortiça seja relevante apenas para os vinhos de eleição; aliás, uma maior quota de cortiça de boa qualidade para rolhamento, tendo em atenção que se trata dum mercado com grande procura e que tende a manter-se como tal, manterá por via da confiança a preferência pela cortiça, sem forçosamente haver uma queda significativa de preços — caso contrário, irá aumentando o risco duma mudança irreversível na orientação do consumo, com a progressiva adopção dos materiais alternativos. Sendo Portugal o mais importante país produtor de cortiça, cabe-nos um papel de grande responsabilidade no acautelamento de tendências negativas do mercado mundial.
Apesar da rentabilidade da exploração da cortiça e do apego de muitos produtores ao sobreiro, a motivação para manterem este uso do solo de que são proprietários não é um dado garantido. Cita-se principalmente a perda irreversível dessa rentabilidade por falta de regeneração dos povoamentos, apesar de poder pensar-se que não é preocupação urgente tendo em conta o longo ciclo de produtividade das árvores, potencialmente de 15 a 20 tiradas de cortiça amadia antes de começarem a entrar em declínio (mas pode ser até menos [Subericultura pp. 162–164]): as árvores que pela idade vão ficando menos produtivas constituem um impedimento para que as árvores jovens se desenvolvam, ou seja, se aquelas não forem abatidas não há regeneração natural, menos ainda por causa do pastoreio que sempre caracterizou a exploração em sistema de montado. Em função disso, Vieira Natividade preconizava uma gestão dos povoamentos orientada para a sustentabilidade da produção, através dum cuidadoso programa de desbastes, enquadrado por inventário minucioso, para conseguir-se uma distribuição uniforme dos sobreiros pelos sucessivos estádios do seu desenvolvimento [Subericultura pp. 217–218, 235]. Tais desbastes seriam ainda um ensejo para a selecção das árvores com melhor potencial de qualidade. Sem dúvida racional, esse modelo de gestão não é possível hoje — o abate não é permitido senão em situações muito estritas — mas não deixa de haver sugestões potencialmente eficazes que passam pela protecção rotativa da regeneração face ao pastoreio, parcela a parcela [Varela et al., 2003b]; e acrescenta-se o problema já antigo das dificuldades para a sobrevivência das árvores jovens, decorrentes da degradação dos solos [Subericultura pp. 124–125]. Talvez por isso, e apontando a crescente decrepitude dos montados de sobro (puros ou mistos) como uma ameaça para o futuro da produção de cortiça, os produtores insistam numa maior liberalização do abate. Tendo em conta a rentabilidade do modelo de exploração dos montados e a manutenção da procura de cortiça de qualidade, essa pretensão não deverá justificar quaisquer receios de vir a ser, pelo menos para a esmagadora maioria dos produtores, um meio de substituir a produção de cortiça por outros usos do solo.
Por outro lado, é de certa forma irónico que o desinteresse dalguns proprietários pela produção de cortiça, quando associado a quebras de rendimento, se prenda com práticas incorrectas que contribuem para a susceptibilidade a doenças ou outros condicionantes da fisiologia das árvores, comprometendo gravemente a quota de cortiça de qualidade extraída [Nogueira 1978]. Este desinteresse encontra-se também entre os que preferem plantações de espécies de crescimento rápido que sejam ecologicamente compatíveis com as áreas actualmente cobertas por sobreiro, já não falando de outros usos do solo mais apetecíveis a curto prazo, e é um factor a ter em conta na evolução do sector.
O insucesso das plantações de sobreiros criados em viveiro também merece muita atenção. Para além da questão de serem mais ou menos correctamente projectadas, e mantidas de acordo com boas práticas, há o problema da adaptação aos locais definitivos, não só pelo facto de tratar-se frequentemente de solos sem passado florestal recente (caso das muito significativas áreas englobadas pela directiva comunitária da Política Agrícola Comum 2080/92), mas também pelo potencial de cada genótipo face às condições disponíveis. A proveniência do germoplasma não tem sido devidamente controlada, sendo provável que muito do material plantado tenha tido origem em regiões muito diferentes, inclusivamente do estrangeiro. O previsível insucesso de muitas plantações representa, para além dos prejuízos económicos directos, mais uma fonte de desmotivação para o cultivo do sobreiro.
2) Nature vs. nurture da qualidade da cortiça
A qualidade da cortiça define-se consensualmente pelo ponto de vista da sua aplicação industrial mais nobre, que é a produção de rolhas para o engarrafamento de vinhos [Vásquez 2002, González-Adrados et al. 2000]. Dentro de margens normais de densidade, dureza e elasticidade (embora esta última só ganhe em ser maximizada), o critério principal para a avaliação das cortiças pelos escolhedores ligados a essa produção é a sua porosidade [Pereira 1998]. A melhor cortiça para rolhas tem uma baixa porosidade que garante a possibilidade de maturação dos vinhos após engarrafamento e a necessária longevidade desse engarrafamento. À medida que as características da cortiça se vão afastando deste padrão, ela é menos valorizada. Natividade [Subericultura pp. 105, 176] exemplificou a distribuição da cortiça tirada em duas parcelas de 1 ha (figura 2.8), que patenteia a grande variação em cada local assim como a proporção minoritária de cortiça de boa qualidade [Ferreira et al. 2000]. Ainda segundo Natividade, de toda a cortiça extraída em Portugal cada ano, apenas 30% tinha a qualidade requerida para a produção de rolhas. Distribuições como as da figura 2.8 levam a concluir que a obtenção de populações produtoras apenas das melhores cortiças teriam de resultar duma forte intensidade de selecção.
 Figura 2.8 — Distribuição da produção de cortiça, em duas
parcelas de 1 ha, por classes de qualidade [Subericultura p.
105]. Trata-se, na parcela 1, dum exemplo de bom arvoredo
em região produtora de boa cortiça, e na parcela 2, dum
exemplo onde apenas houve desbastes selectivos visando
reduzir a proporção de cortiças de má qualidade.
Figura 2.8 — Distribuição da produção de cortiça, em duas
parcelas de 1 ha, por classes de qualidade [Subericultura p.
105]. Trata-se, na parcela 1, dum exemplo de bom arvoredo
em região produtora de boa cortiça, e na parcela 2, dum
exemplo onde apenas houve desbastes selectivos visando
reduzir a proporção de cortiças de má qualidade.Embora os desbastes, que em todo o caso serviam também para uma gestão racional dos povoamentos, sejam uma forma de selecção [Subericultura pp. 104, 236], não se pode considerar que houvesse alguma vez melhoramento genético para a qualidade da cortiça. Aliás, logo que se tornou possível o aproveitamento integral da produção (incluindo o refugo e as aparas) para a indústria de aglomerados, associado a um declínio dos preços, mesmo os esforços de selecção por via dos desbastes foram interrompidos.
A porosidade aumenta com o volume ocupado pelos canais lenticulares da cortiça, o qual por sua vez depende do número de lentículas por unidade de área de entrecasco e do calibre dos canais [Subericultura p. 179, González-Adrados et al. 2000, Vásquez 2002]. Apenas o primeiro destes factores (isto é, o arranjo lenticular do entrecasco) parece ter uma forte base genética, pelo menos a julgar pela sua relativa constância em cada árvore, independentemente da altura no tronco, da idade (e/ou número de tiradas anteriores), etc.. As lentículas diferenciam-se, durante a primeira formação do câmbio suberofelodérmico, sob câmaras estomáticas na epiderme do ramo jovem [Subericultura p. 99]. Para obviar à espera de 35-40 anos até extrair-se a cortiça secundeira, que é quando em definitivo se avalia o valor duma árvore em termos de qualidade da cortiça, pode fazer-se uma avaliação precoce utilizando análise de imagem em cortes transversais de ramos jovens: um maior desenvolvimento do entrecasco nos de mais de 1 ano, e da cortiça virgem nos de diâmetro inferior a 1 cm, correlaciona-se com maior porosidade [Vásquez 2002]. A espessura do entrecasco ou, mais precisamente, do floema inactivo, determina a profundidade onde a partir deste se regenera o felogénio traumático, após descortiçamento, o que para providenciar o necessário arejamento requer lentículas de diâmetro maior, embora a superfície de contacto com os raios medulares do lenho, também variável entre árvores, contribua para facilitar as trocas gasosas [Vásquez 2002].
Muitas cortiças da melhor qualidade têm numerosos canais lenticulares mas de reduzido diâmetro, resultando na sua baixa porosidade, por isso não se atribui a este factor uma relevância determinante da porosidade da cortiça [Subericultura pp. 164–165], quando comparada com factores ambientais como por exemplo os que influenciam a intensidade de crescimento da cortiça: as temperaturas, a mobilização do solo, a captação da luz e da água, e de nutrientes do solo, o parasitismo, etc.. Não são as condições mais propícias à actividade vegetativa do sobreiro [Natividade 1939], nomeadamente a disponibilidade hídrica [Vásquez 2002], as que coincidem com as da cortiça de melhor qualidade, daí que as regiões mais expostas à influência marítima não produzam cortiças de 1ª e 2ª qualidade, e que nas regiões montanhosas com povoamentos mais densos, onde o crescimento da cortiça é menos intenso, a porosidade tenda a ser menor [Subericultura p. 179]. Em Portugal, uma grande parte dos povoamentos com cortiça de melhor qualidade situam-se nas regiões de contacto com as azinheiras, e embora pareça haver uma correlação negativa entre essa qualidade e as condições mais propícias à actividade vegetativa do sobreiro, é uma questão ainda por resolver [Vásquez 2002].
A reduzida porosidade das melhores cortiças não as faz substancialmente mais densas, aliás, é no refugo que as cortiças mais densas, e concomitantemente duras, são colocadas. Aparte os casos em que os anéis suberosos depositados anualmente são demasiado delgados e com paredes celulares espessas, o que em Portugal apenas se encontraria em sobreiros muito velhos [Subericultura p. 181], a densidade elevada resulta de inclusões lenhificadas que caracterizam as cortiças preguentas ou madeirentas, cujo desenvolvimento é anómalo e poderá atribuir-se a factores genéticos, mas através de processos não relacionáveis com as causas de porosidade (cf. secção 3).
Para além das tendências regionais já referidas, que se relacionam com as condições edafoclimáticas e os modelos de exploração que as caracterizam, grande parte da variação na qualidade da cortiça é local, ou seja, em todos os povoamentos se produz cortiça de qualidades muito diversas (figura 2.8 [Ferreira et al. 2000]). Tanta variação local não parece explicar-se apenas por factores ambientais, mas o facto de observar-se nas descendências de sobreiro uma grande heterogeneidade fenotípica, a par da alogamia que caracteriza o género Quercus, leva a atribuir-se aos sobreiros, como aliás aos carvalhos em geral, uma elevada heterozigose [Subericultura p. 104] — ou seja, mesmo no cruzamento entre dois sobreiros genotipicamente superiores, podem facilmente produzir-se descendentes de má produção. A relação entre este quadro geral, em que muita da presumível variância genotípica será não-aditiva, e a variação na qualidade da cortiça em particular, é totalmente desconhecida.
Em suma, embora seja convicção generalizada que existe uma componente genética que contribui para a variação na qualidade da cortiça, a mesma ainda permanece por avaliar com ensaios de proveniência e de descendência adequados. Apenas muito recentemente houve um esforço, a nível internacional, de estabelecer esses ensaios para acompanhamento no futuro [Varela 2001, Varela et al. 2003b, Almeida et al. 2005, http://europa.eu.int/comm/research/agro/fair/en/pt0202.html], portanto e aparte a possibilidade de avaliar precocemente a qualidade da cortiça, não será antes de algumas décadas que talvez se conheçam os resultados. No presente contexto, importa principalmente considerar a possível implicação do hibridismo na qualidade da cortiça.
3) Cortiça preguenta e a hipótese de sua origem nos híbridos com azinheira
Vieira Natividade, utilizando uma técnica por si desenvolvida com sobreiro para a análise histológica das peridermes (isto é, dos derivados do câmbio suberofelodérmico), pela qual pôde analisar a relação entre a actividade do câmbio suberofelodérmico e a anatomia microscópica da cortiça amadia daí resultante [Natividade 1934], fez uma comparação entre o que seriam os padrões histológicos característicos da azinheira, de sobreiro e dos híbridos de sobreiro e azinheira [Natividade 1936]. Estes últimos apresentavam uma intercalação de ambos os padrões histológicos (de sobreiro e de azinheira), alternando entre si de modo variável, consoante a árvore e mesmo dependendo da zona do tronco. Mais precisamente, enquanto nos sobreiros a actividade do câmbio suberofelodérmico, produzindo peridermes contínuas, é persistente durante longo tempo — não sendo substituído senão como consequência do descortiçamento — na azinheira essa actividade é de curta duração, diferenciando-se novos núcleos de tecido gerador, limitados em extensão e englobando nos intervalos restos de líber, numa sucessão periclinal de inúmeras formações de consistência lenhosa, que constituem o ritidoma; nos híbridos, apesar da existência de peridermes contínuas como nos sobreiros, intercalam-se entre elas regiões características da azinheira [Natividade 1936]. Deste modo se explicaria a aparência intermédia destes híbridos em termos de cobertura do tronco [Natividade 1936] (cf. fotografias no Apêndice I secção C2).
O padrão observado nos híbridos foi interpretado como análogo, embora em diferente grau, ao do que anteriormente havia sido observado em cortiça “preguenta” ou “madeirenta” [Natividade 1934, fig. 67, pp. 89–96]. Este tipo de cortiça é considerada de muito má qualidade porque é atravessada por veios lenhosos que dificultam a sua extracção e o seu processamento industrial, e as imagens publicadas no conjunto destes dois estudos denotam, na interpretação de Natividade, o mesmo fenómeno: a intercalação de tecido lenhoso (peridermes parciais, dispostas em arcos sobrepostos) que interrompem a continuidade do tecido suberoso característico da cortiça normal (camadas contínuas, paralelas ao câmbio). Para Natividade, os sobreiros de cortiça preguenta originam-se da descendência de híbridos, e por terem genes de azinheira introgredidos produzem este tipo particular de cortiça. Por outras palavras, mesmo que os híbridos sejam relativamente raros, a prevalência de genes de azinheira nos sobreiros pode ser importante, contribuindo para uma parte da cortiça de refugo.
Um sobreiro de cortiça preguenta é facilmente identificado desde as primeiras tiradas, pois o seu entrecasco se mostra muito enrugado, com projecções em relevo pontiagudas, alinhadas longitudinalmente segundo a orientação dos feixes liberinos (cf. Apêndice I secção C3). A ser verdade que estes sobreiros têm origem híbrida, seria talvez legítimo erradicá-los, o que não ofereceria dificuldades de maior visto que os tiradores os sabem reconhecer; essa erradicação não foi levada a cabo naquela época, não talvez por eventuais dúvidas sobre a hipótese avançada por Natividade, mas porque essa cortiça, apesar da sua má qualidade e da dificuldade em ser tirada, é sempre uma minoria da que se extrai e pode ser aproveitada para a indústria de aglomerados [Subericultura p. 235].
Contudo, se no futuro a percepção da cortiça preguenta mudar, não serão apenas esses sobreiros
(e os híbridos) que podem tornar-se alvo de abate selectivo: as azinheiras dos montados mistos acabam
por ser consideradas uma ameaça à “pureza” dos sobreiros nesses montados (conceito dúbio, como se
verá), o que teria consequências muito mais graves, inclusivamente a nível ecológico. Por isso a pressão
que já exista ou possa vir a desenvolver-se no sentido da eliminação das azinheiras dos montados mistos
não parece corresponder a uma real necessidade de proteger a qualidade da cortiça, mas antes a uma
ânsia de expandir a cultura do sobreiro à custa da eliminação das azinheiras. A cultura do sobreiro em
associações naturais, biologicamente mais equilibradas e com defesas mais eficientes (citando-se
associações com o medronheiro, o carrasco e o zambujo, e implicitamente a azinheira [Subericultura p.
219], referência 9330 do Plano Sectorial da Rede Natura 2000
![]() ), é uma vantagem que talvez não chegue
a ser anulada pelo risco de hibridismo, no caso com a azinheira, sobretudo se esse hibridismo for
suficientemente raro. Sem haver uma medida para a taxa de hibridismo nos montados mistos de sobro
e azinho, a consideração por parte de Vieira Natividade de que constituem uma associação pouco
prudente (mesmo que não se aproveite a regeneração natural [Subericultura p. 219]) não é sustentável
[Varela 1995].
), é uma vantagem que talvez não chegue
a ser anulada pelo risco de hibridismo, no caso com a azinheira, sobretudo se esse hibridismo for
suficientemente raro. Sem haver uma medida para a taxa de hibridismo nos montados mistos de sobro
e azinho, a consideração por parte de Vieira Natividade de que constituem uma associação pouco
prudente (mesmo que não se aproveite a regeneração natural [Subericultura p. 219]) não é sustentável
[Varela 1995].
A hipótese sobre a origem dos sobreiros de cortiça preguenta, formulada por Natividade, é totalmente concordante com a evidência por ele apresentada, mas assenta no pressuposto de que as inclusões de tecido lenhoso nessa cortiça têm uma causa diversa doutras que esse mesmo autor descreve devidas a formações de esclerênquima, em feixes liberinos de maior calibre ou sob as lentículas, que também se evidenciam por relevos característicos no entrecasco sem forçosamente estarem associados a cortiça de má qualidade [Subericultura pp. 152–3]. As comparações feitas com híbridos e azinheiras [Natividade 1936] usaram como termo de referência da espécie “pura” de sobreiro as árvores que mais fielmente reproduziam o tipo específico [Natividade 1934 p. 128], enquanto uma comparação mais sistemática da variação em sobreiros de cortiça não-preguenta, das mais diversas qualidades, especialmente se acompanhada por caracterização através de marcadores genéticos dessa qualidade, poderia permitir uma melhor avaliação sobre a distinção dos sobreiros de cortiça preguenta em relação à generalidade dos restantes. Formalmente, nunca foi determinado se a causalidade da cortiça preguenta é genética, ambiental, ou uma interacção entre ambas. Com estas lacunas de informação, não é lícito tirar conclusões sobre a hipótese de Natividade, embora se deva levar em conta os seus fortes argumentos, com que permanece muito plausível.
Os híbridos entre sobreiro e azinheira, que na nomenclatura botânica são designados Quercus ×
morisii Borzi, embora seja preferível a designação Q. × morisii f. mixta Franco & Vasc. por implicar
apenas a subespécie de azinheira rotundifolia [Vasconcelos & Franco 1954], são bem conhecidos das
populações alentejanas, que muitas vezes lhes reconhecem o carácter híbrido. Além disso, utilizam a
casca na preparação dum chá a que atribuem propriedades abortivas, como é visível pelas mutilações que
a mesma apresenta em diversas árvores ([Natividade 1936]; cf. Apêndice I secção C2). É aliás pela casca
que mais facilmente se identificam estas árvores no campo, quando já adultas, pois pela sua consistência
não se confunde com a cortiça dos sobreiros, e pela espessura e cor não se confunde com o ritidoma da
azinheira. O nome popular que se lhe atribui actualmente é carvalho cerqueiro (ou apenas cerqueiro)
referido por Vieira Natividade [1936], ao que parece distinto do “negral”, nome usado pelo menos no
Alto Alentejo para referir formas relativamente frequentes que são sistematicamente eliminadas pelos
produtores durante as desmoitas, pela presunção de serem híbridos; estas duas designações aparecem na
língua portuguesa também para Q. pyrenaica, e regista-se cerqueiro-bravo para Q. lusitanica [Franco
1990, Vasconcelos & Franco 1954]
![]() . O presente estudo não incluiu os “negrais”.
. O presente estudo não incluiu os “negrais”.
Deve-se a Pereira Coutinho a primeira referência detalhada e sistematizada aos híbridos entre sobreiro e azinheira em Portugal [Coutinho 1888], para os quais registou os nomes de carvalho cerqueiro, carvalho de sequeiro e azinheira macha. Define-os como híbridos em função de critérios de aplicação geral, discutidos em detalhe nesse mesmo trabalho, incluindo o apresentarem caracteres importantes de uma ou outra das duas espécies a que se lhes atribui a origem, o aparecerem predominantemente na proximidade de ambas e, apesar de considerá-los frequentes, encontrarem-se dispersos (este último critério, a que o autor atribuía grande importância, pode talvez não aplicar-se em muitos casos de hibridismo entre Quercus, pelo menos nas espécies americanas [Benson et al. 1967, Williams et al. 2001, Gonzalez-Rodriguez et al. 2004a, b]). Acrescenta que o facto de serem férteis não é excepção dentro do género Quercus. Para a diagnose dos híbridos, sugere nas chaves dicotómicas, entre outros caracteres, o revestimento do tronco (que não considera suberoso), a pilosidade das anteras (glabras como na azinheira), o ângulo no ponto de inserção das nervuras secundárias na principal (relativamente agudo, como no sobreiro) e a disposição das escamas da cúpula (tendencialmente compridas e soltas, como no sobreiro). Num suplemento à Flora de Portugal, Coutinho parece basear-se no trabalho então ainda por publicar de Vieira Natividade [Natividade 1936] para considerar que o revestimento do tronco se compõe de tecido suberoso muito mais desenvolvido que na azinheira [Coutinho 1935].
Vasconcelos e Franco [1954] referenciam os cerqueiros em 14 concelhos, a maior parte em Trás-os-Montes e Alto Douro e no Alto Alentejo, mas abrangendo também o Baixo Alentejo, Olhão e mesmo Loures. A diagnose é feita através das folhas, afins às do sobreiro pelo ângulo de inserção das nervuras secundárias, e pela nitidez dessas nervuras na página superior (mas que no tipo juvenil se aproximam da azinheira pelo recorte espinhoso-dentado das folhas e pelo ângulo de inserção das nervuras secundárias na principal, menos agudo, e no tipo adulto pelos pecíolos mais curtos); e pelo tronco, onde consideram não haver revestimento suberoso mas ritidoma. Os nomes vernáculos cerqueiro e azinheira macha são listados para este híbrido.
Dispondo das descrições para os esquemas da Flora Europæa e de Krüssmann (figura 2.1), podem acrescentar-se caracteres diagnosticantes a priori entre o sobreiro e a azinheira (tabela 2.2), a maior parte deles associados às flores e frutos.
Tabela 2.2 — Critérios diagnosticantes entre sobreiro e azinheira em função dos esquemas taxonómicos delineados na figura 2.1 [Tutin 1964, Franco 1990, Krüssmann 1978].
Flora Europæa |
Esquema de Krüssmann |
||
Subgénero Cerris |
Subg. Sclerophyllodrys |
Secção Suber |
Secção Ilex |
1. Folhas persistentes ou caducas, denticuladas a penatifídeas 2. Anteras geralmente obtusas, pilosas 3. Perianto das flores femininas pateniforme, dividido em lóbulos lineares 4. Estiletes enrolados 5. Frutos de maturação bienal, mais raramente anual 6. Endocarpo glabro |
1. Folhas persistentes coriáceas, na fase juvenil frequentemente dentadas ou asserado-espinhosas 2. Anteras geralmente mucronadas, glabras ou pilosas 3. Perianto das flores femininas campanulado, lóbulos obtusos 4. Estiletes lineares 5. Frutos de maturação anual ou bienal 6. Endocarpo tomentoso |
1. Estilete linear, bastante pontiagudo, direito ou deflexo 2. Escamas da cúpula livres ou só ligeiramente aplicadas 3. Frutos amadurecem em 1 ou 2 anos |
1. Estilete curto, arredondado 2. Escamas da cúpula aplicadas 3. Frutos amadurecem em 1 ano 4. Folhas espessas, coriáceas |
Alguns destes caracteres (perianto feminino e estiletes, maturação dos frutos) não se verificam claramente, mesmo no sobreiro e na azinheira, nas descrições de Vasconcelos e Franco [1954]. Nestas confirma-se que os híbridos têm folhas juvenis semelhantes às de azinheira (espinhoso-dentadas), anteras pubescentes como o sobreiro (em contradição a Coutinho [1888]); acrescente-se a floração em Abril, isto é, entre a da azinheira (Março-Abril) e a do sobreiro (Maio), os gomos pubescentes como em sobreiro, e ráquis dos amentilhos hirsutos como na azinheira.
No estudo sobre as peridermes dos híbridos [Natividade 1936] incluem-se observações sobre outros aspectos destas árvores, donde se destacam a referência a trabalhos que reiteradamente registam a sua presença na Andaluzia, uma análise detalhada da consistência das folhas, e a expressão fenotípica nas suas descendências.
Quanto às folhas, Vieira Natividade confirmou a aparência predominantemente de sobreiro (forma, recorte, nervação lateral), bem como a cor do tomento na página inferior semelhante à de azinheira, já indicada por Coutinho [1888] e mais tarde reiterada por Vasconcelos e Franco [1954], mas sobre este carácter aparentemente constante definiu-o como difícil de apreciar com rigor (o mesmo considerou sobre o brilho da folha na página superior, semelhante ao da azinheira). Junta-lhes a consistência coriácea das folhas na fase adulta, associada a uma espessura semelhante à de azinheira (superior à do sobreiro), resultante das camadas hipodérmicas, praticamente inexistentes em sobreiro, e da espessura das paredes celulares em contacto com a cutícula.
Quanto ao revestimento do tronco, apresenta-se nas árvores mais desenvolvidas
![]() «uma grande
analogia com o sobreiro pelo enguiado, a cor, a espessura e a consistência predominantemente suberosa»
[Natividade 1936]. Ainda assim, e também pela arquitectura anatómica que descreveu (cf. secção B3),
o fenótipo destas árvores era intermédio entre as duas espécies progenitoras.
«uma grande
analogia com o sobreiro pelo enguiado, a cor, a espessura e a consistência predominantemente suberosa»
[Natividade 1936]. Ainda assim, e também pela arquitectura anatómica que descreveu (cf. secção B3),
o fenótipo destas árvores era intermédio entre as duas espécies progenitoras.
Tratando-se de cerqueiros disseminados em povoamentos de resto “puros” de sobro, em Grândola e Mora, e tendo em conta a analogia com a fertilidade dalguns sobreiros com flores masculinas estéreis observados anteriormente, as descendências destes híbridos deveriam descender em grande parte da polinização com pólen de sobreiro. No entanto, duma mesma árvore, observavam-se plantas juvenis indistintas das azinheiras nessa fase, se bem que minoritárias, algumas intermédias, e a maior parte indistinguíveis dos sobreiros. Este resultado, análogo ao que se obteve de híbridos Q. × hispanica e Q. × airensis, serviu para atestar a origem híbrida desses cerqueiros.
Finalmente, os cerqueiros apresentavam uma intensa esclerificação dos raios medulares largos, que é característica dos sobreiros mas não das azinheiras [Natividade 1936].
3) Outras observações e súmula
As emissões de isoprenóides pelas folhas dos Quercus, que derivam do Carbono recém-fixado pela fotossíntese e se julga terem um papel estabilizador das membranas face ao calor, atenuando a inibição da fotossíntese pelas altas temperaturas [Loreto et al. 1998, Delfine et al. 2000], consiste de isoprenos na generalidade do género, de monoterpenos nas azinheiras (e em Q. coccifera) e, ainda em sobreiro, apesar de neste (e em Q. cerris) se ter afirmado a ausência destas emissões [Loreto et al. 1998]. A emissão de monoterpenos pelos sobreiros [Staudt et al. 2004, Pio et al. 2005] retira-lhe o carácter diagnosticante em relação à azinheira e, presume-se, aos híbridos. A codominância observada nos híbridos entre azinheira e Q. robur (Q. × turneri), e entre azinheira e Q. canariensis (?), isto é, um padrão misto de monoterpenos e isoprenos [Schnitzler et al. 2004; Staudt et al. 2004], não pode servir para interpretar a emissão em sobreiro como um indicador de pressupostas introgressões de genótipo de azinheira [Staudt et al. 2004], principalmente por não excluir-se a possibilidade das variações observadas entre povoamentos serem de causa ambiental.
Ao contrário da subespécie ilex, a subespécie rotundifolia apresentou valores sempre muito próximos dos de sobreiro nos parâmetros das curvas de pressão-volume, que no contexto mais alargado de definir grupos funcionais dentro dos Quercus mediterrânicos permitem inferir a fisiologia do aproveitamento da água disponível; globalmente, este estudo definiu um grupo perenifólio/ esclerófilo incluindo os três táxones junto com Q. coccifera e Q. chrysolepis Liebm. [Corcuera et al. 2002]. Este resultado sugere que, em termos de aproveitamento da água, a fisiologia dos híbridos entre sobreiro e azinheira deverá ser semelhante à das duas espécies respectivas.
A composição das sementes em compostos fenólicos, derivados do ácido gálico [Cantos et al. 2003] mostra predominância de espectros do tipo ácido gálico na azinheira, e dos do tipo ácido elágico no sobreiro. Este último também se distinguiu pela abundância de dilactona de ácido valoneico no tegumento da semente. Nos híbridos não foram feitas observações.
No decurso do presente trabalho houve a oportunidade de observar diversas árvores, confirmando-se as descrições existentes e acrescentando-se alguns caracteres:
Analogamente às azinheiras, estes híbridos são muito tardios na diferenciação da cobertura do tronco. Em consequência, nem todos os híbridos observados no campo apresentam a suberificação que lhes é característica na maturidade (cf. Apêndice I secção C2), e é possível que a insistência dalguns taxonomistas em descrever a casca como coberta por ritidoma não suberoso se relacione com este desfasamento. A semelhança das folhas na fase juvenil com as da azinheira nessa mesma fase, talvez uma característica materna se se considerar provável a tendência para as azinheiras serem o progenitor feminino dos híbridos (cf. parte II secções C3 e D2), deve levar a confundir com as azinheiras as plantas híbridas, que só ganham caracteres de sobreiro (revestimento suberoso, folhagem, caracteres florais) à medida que se tornam adultas. Isto talvez explique porque os híbridos nem sempre são detectados nos povoamentos mistos, a tempo da desmoita. Tal como no trabalho de Natividade [1936], observaram-se plantas descendentes dos híbridos semelhantes às de azinheira como ao sobreiro.
Algumas das árvores mais maduras (ritidoma suberoso) tinham envergadura e altura não inferior ao que é usual observar nas espécies progenitoras (cf. Apêndice I secção C2).
As folhas conferem à copa a tonalidade de sobreiro, isto é, um verde não acinzentado (este carácter é mais difícil de observar na Primavera, por causa da nova folhagem e do candeio). Confirmou-se a observação de Natividade [1936] que a espessura, em corte transversal, é a das folhas de azinheira e não a das de sobreiro.
Mostrou-se impraticável utilizar na diagnose a cor do tomento na página inferior das folhas (como previu Natividade [1936]), bem como a da textura mais ou menos acetinada do interior da cúpula do fruto [Coutinho 1888, Vasconcelos & Franco 1954].
A comparação das escamas da cúpula dos frutos dos híbridos com as de azinheira ou de sobreiro não foi conclusiva, embora não pareça ser diagnosticante, face à grande variação neste carácter em sobreiro [Coutinho 1888, Natividade 1950, Vasconcelos & Franco 1954, Franco 1990].
Confirmou-se ainda que a floração da azinheira é mais precoce que a do sobreiro, em média de 4 a 6 semanas [Varela 1995, Brás 1999, Varela et al., em preparação], e com base em observações no ano de 2003 os híbridos distribuem-se no período intermédio entre as duas espécies (cf. Apêndice II secção E1).
Os grãos de pólen dos híbridos (cf. Apêndice II secção E2) têm dimensões bastante próximas das dos provenientes de azinheira, embora se tivesse notado uma maior dispersão de valores; outra observação de interesse foi a taxa elevada de grãos de pólen “incompletos”, geralmente reduzidos à exina, o que parece ser um indicador de abortamento, característico dos híbridos de Quercus [Rushton 1993].
Reunindo a informação aqui incluída, deduz-se um quadro da expressão nos híbridos dos caracteres morfológicos e fenológicos das espécies progenitoras (tabela 2.3):
Tabela 2.3 — Expressão de alguns caracteres discriminantes entre sobreiro e azinheira nos híbridos Quercus × morisii f. mixta. Para cada órgão, separam-se os caracteres semelhantes à azinheira, ao sobreiro, ou intermédios. As chamadas numeradas referem as fontes bibliográficas citadas em rodapé.
Órgão |
Expressão nos híbridos |
Tronco |
azinheira: desenvolvimento tardio do ritidoma intermédio: suber atípico no adulto1 sobreiro: intensa esclerificação dos canais medulares largos1 |
Folhas |
azinheira: fase juvenil de recorte espinhoso-dentado2; ângulo de inserção das nervuras secundárias2; comprimento do pecíolo2 e espessura do limbo1 na fase adulta; cor do tomento na página inferior1,2,3 sobreiro: gomos pubescentes2, folhas do adulto de recorte denticulado2, nervuras secundárias visíveis na página superior e inseridas em ângulo agudo1,2,3, cor da copa |
Floração masculina |
azinheira: ráquis dos amentilhos hirsutos2, dimensão média dos grãos de pólen intermédio: época de floração2 sobreiro: anteras pubescentes2 |
Frutos |
intermédio: textura do interior da cúpula2,3 sobreiro: escamas da cúpula2,3 |
1 [Natividade 1936] 2 [Vasconcelos & Franco 1954] 3 [Coutinho 1888]
Dois estudos de Elena-Rossellò et al. [1992, 1997] sugeriram que algumas das variantes isoenzimáticas observadas em sobreiro e azinheira eram discriminantes entre as duas espécies, e com base nesse pressuposto deduziram que existe hibridismo nas populações estudadas. O primeiro estudo [Elena-Rossellò et al.1992], citado frequentemente, relata a ocorrência de 1 híbrido entre 35 indivíduos jovens, num povoamento misto em Cáceres maioritariamente com azinheiras; mas o critério de discriminação baseou-se em apenas 50 azinheiras e 41 sobreiros que serviram de “padrão”, enquanto as amostragens muito mais extensas de Michaud et al. [1995] em azinheira e de Toumi & Lumaret [1998] em sobreiro vieram a comprovar que os alelos considerados “discriminantes” nos loci Lap1 e Acph1 não o eram de todo; e no estudo de 1997, o “alelo” 2 em Est1 ocorria em ambas as espécies, assim invalidando a conclusão inicial. Na melhor das hipóteses, o único caso possivelmente envolvendo hibridismo nestes estudos é uma azinheira adulta com o alelo 3 de sobreiro em Est1 [Elena-Rossellò et al. 1997], mas este locus não foi escrutinado em amostragens mais representativas (excepto num trabalho só em sobreiro, mas onde a alteração da nomenclatura dos zimogramas não permite uma comparação [Elena-Rossellò et al. 1996]), de modo que, com amostragens tão reduzidas, é questionável se se pode interpretar dessa maneira este marcador. Aparentemente, os autores do primeiro estudo consideraram que cada banda presente em ambas as espécies é indício de fluxo genético interespecífico, sem chegarem a levar em conta as possibilidades de homoplasia ou de polimorfismo partilhado.
Toumi e Lumaret [1998] detectaram em sobreiros alelos minoritários que afirmam serem de azinheira. Tendo em conta definirem como alelos de sobreiro apenas os que não fossem encontrados noutras espécies (azinheira, carrasco), há lugar a perguntar como é que as amostragens de apenas algumas dezenas de indivíduos por povoamento iriam esgotar a diversidade intra-específica de sobreiro... note-se que tal definição levou a que em 7 loci fossem identificados 25 alelos “de azinheira ou de carrasco” introgredidos em sobreiro! Em apoio à sua interpretação, os autores citam duas árvores na Catalunha com morfologia intermédia que eram heterozigóticas para alelos “discriminantes” (não especificados) em 2 loci, mas também ilustram a completa resolução das duas espécies num povoamento misto da Sicília, usando os genótipos em todos os loci, apesar de considerarem haver alelos de azinheira em alguns sobreiros (ou talvez o contrário, cf. final deste parágrafo). Apesar do critério usado parecer muito arbitrário, da utilização de diferenças interespecíficas demasiado subtis para constituirem verdadeiros marcadores discriminantes (por exemplo Lap1.96 em azinheira e Lap1.97 em sobreiro) e da prevalência de sobreiros supostamente introgredidos ser dramaticamente elevada em alguns locais, não deixa de haver resultados que lhe parecem dar alguma plausibilidade [Michaud et al. 1995, Toumi & Lumaret 1998]: no locus Pgi–1, o alelo “ilex” mais comum em sobreiro era o maioritário em azinheira em toda a distribuição desta última, e a generalidade dos outros alelos “ilex” ocorrendo em sobreiros eram comuns em toda a distribuição da azinheira ou pelo menos ocorriam nas duas espécies nas mesmas regiões. Os autores do estudo em sobreiro não ilustraram com o mesmo detalhe outros loci, o que poderia não só reforçar as suas interpretações e permitir uma mais completa cartografia da postulada introgressão. Com base apenas no locus Pgi–1, haveria introgressão de genes de azinheira em sobreiros de Marrocos (planalto central e Bab Azhar), França (Languedoque) e Espanha (Catalunha central, Serra Morena Ocidental, Sul da Andaluzia), e de genes de sobreiro em azinheiras da Sicília.
Belahbib et al. [2001] recorreram a diversos marcadores de DNA citoplásmico (plastidial e mitocondrial) para descreverem 6 haplótipos em sobreiros e azinheiras de Marrocos (PCR-RFLP [Demesure et al. 1995, Dumolin-Lapègue et al. 1997b]). O haplótipo mais frequente em azinheira era o mesmo que predominava nas amostragens de sobreiros do Alto Atlas, parte do planalto central, e em Bab Azhar (Atlas Médio), formando uma mancha geográfica parapátrica em relação às restantes amostragens de sobreiro, onde normalmente este haplótipo não era detectado. Em menor escala, o haplótipo de sobreiro mais frequente observava-se em amostras de azinheira de duas áreas não-contíguas dominadas por sobreiro. A interpretação destes resultados invocou a mediação de híbridos naquilo que já tinha sido descrito para Q. robur/Q. petræa [Petit et al. 1997, Belahbib et al. 2001], como discutido na parte II, secção D2.
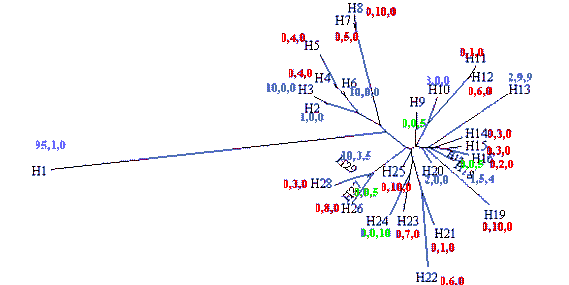
Collada et al. [2002] usaram a mesma estratégia para caracterizarem amostras recolhidas na Península Ibérica de sobreiro, azinheira e carrasco, mas incluindo dois novos marcadores plastidiais (e abdicando do aparentemente redundante marcador mitocondrial [Belahbib et al. 2001]) que lhes permitiram diferenciar 29 haplótipos diferentes, dos quais apenas um foi classificado como “suber” por ser o mais comum em sobreiro e por causa do radical distanciamento no dendrograma do conjunto (H1 na figura 2.9a; utilizando informação detalhada posteriormente [Jiménez et al. 2004], seria o mesmo “suber” S1 identificado em Marrocos [Belahbib et al. 2001, Lumaret et al. 2005]). Para os restantes haplótipos, não há correspondência entre a posição no dendrograma e as espécies em que ocorrem, e a presença dalguns haplótipos em duas ou mesmo nas três espécies, assim como a sua colocalização em 3 das 14 populações (figura 2.9b), não corresponde a um padrão geográfico tão claro como em Marrocos.
Mas há uma esclarecedora correspondência entre distribuição geográfica e parentesco dos haplótipos “ilex-coccifera”, independentemente da espécie onde ocorrem (figura 2.9a,b): os ramais do dendrograma H26–H29 no litoral de Portugal, H2–H6 e H11–H12 na costa mediterrânica (se se considerar H2 introduzido na Andaluzia a partir dessa proveniência, o que é plausível) e H13–H20 na metade Sul de Espanha e no Alentejo. A segregação geográfica entre ramais corresponderia a um efeito fundador antigo, talvez relacionado com o recuo das espécies na última fase glaciar, e a diferenciação de haplótipos dentro de cada ramal resultaria de divergências evolutivas mais recentes, nalguns casos associadas a fixação/perda de haplótipos (por exemplo, H27 em carrasco, H26 e H28 em azinheira; mas seria fortemente desejável que o número de árvores observadas por espécie e por local fosse maior, para caracterizar os polimorfismos existentes, visto que cada haplótipo, à excepção de H1, está representado por quando muito 10 indivíduos de cada espécie). Vista desta maneira, a variabilidade existente não implica forçosamente a invasão (pelo sobreiro) dum habitat já ocupado por uma das espécies (sobretudo a azinheira, neste caso), por hibridismo com essa espécie seguido de “submersão polínica” dos híbridos e linhagens deles descendentes [Petit et al. 1997, Belahbib et al. 2001]: é mais provável que os refúgios na última fase glaciar tenham servido para uma fixação dos haplótipos percursores de diferentes ramais “ilex-coccifera” (H2–H29), associada a uma prolongada oportunidade de hibridação entre as 3 espécies e por isso à formação dum fundo genético comum, localmente. Apenas nos refúgios percursores das populações de sobreiro ocupando hoje a faixa Sul-Norte central se teria fixado H1 (figura 2.9b). A partilha de marcadores AFLP (nucleares) entre três espécies da secção Stenocarpæ levou a formular-se uma hipótese análoga [Kashani & Dodd 2002], aparentemente perfilhada pelos autores dum estudo ibérico mais recentemente [Valbuena-Carabaña et al. 2005].
a)
 b)
b)
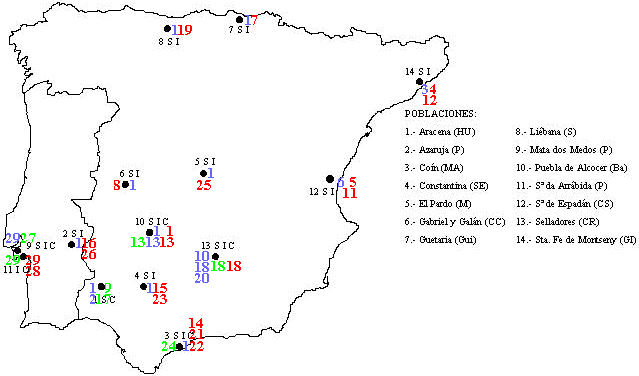 Figura 2.9 — Distribuição dos 29 haplótipos caracterizados por Collada et al. [2002], adaptado das
figuras desse estudo. a) Dendrograma com os graus de semelhança entre haplótipos (H1–H29). Para
cada um indica-se também o número de indivíduos observado de cada espécie (por ordem, Q.suber,
Q. ilex, Q. coccifera), usando-se um código de cores para facilitar a identificação de cada espécie
predominante: azul, Q. suber; verde, Q. coccifera; vermelho, Q. ilex. b) Distribuição geográfica dos
locais de colheita, à qual foram sobrepostas as ocorrências dos haplótipos (1–29), com o mesmo
código de cores a indicar cada espécie.
Figura 2.9 — Distribuição dos 29 haplótipos caracterizados por Collada et al. [2002], adaptado das
figuras desse estudo. a) Dendrograma com os graus de semelhança entre haplótipos (H1–H29). Para
cada um indica-se também o número de indivíduos observado de cada espécie (por ordem, Q.suber,
Q. ilex, Q. coccifera), usando-se um código de cores para facilitar a identificação de cada espécie
predominante: azul, Q. suber; verde, Q. coccifera; vermelho, Q. ilex. b) Distribuição geográfica dos
locais de colheita, à qual foram sobrepostas as ocorrências dos haplótipos (1–29), com o mesmo
código de cores a indicar cada espécie.
Jiménez et al. [2004], no que parece ser uma extensão do trabalho de Collada et al. [2002], identificaram nada menos que 81 haplótipos diferentes, aparentemente formando três ramais bem diferenciados, um deles exclusivo de sobreiro (“suber”), outro partilhado entre azinheira e carrasco (“ilex-coccifera II”), e outro representado nas 3 espécies (“ilex-coccifera I”, o mais diversificado). Do ponto de vista do sobreiro, 72% dos indivíduos eram portadores de um de 4 haplótipos “suber”, sendo os restantes representantes de 23 haplótipos “ilex-coccifera”, 16% com um haplótipo presente nas 3 espécies, 7% com um partilhado só com azinheira, 0,35% com um partilhado só com carrasco, e 4,42% aparentemente exclusivos de sobreiro, apesar de serem “ilex-coccifera I”. Espera-se que a informação sobre a distribuição geográfica de tão complexa variabilidade seja publicada, para verificar se a hipótese acima proposta para os dados de Collada et al. [2002] se mantém plausível.
A amostragem de sobreiros em toda a extensão de distribuição do sobreiro veio revelar padrões de distribuição filogeográfica bastante concordantes [Lumaret et al. 2005]. Sem abdicarem da abordagem PCR-RFLP mas baseando o trabalho em RFLP do DNA plastidial, identificaram só com esta técnica 8 haplótipos “suber”, apesar da amostragem ser muito menor (daí que talvez só 6 haplótipos “ilex-coccifera” fossem detectados). Neste trabalho é notável a coincidência entre o parentesco de 7 desses haplótipos e a sua distribuição geográfica diferenciada no sentido Leste-Oeste, com o ramal S1-S2 do dendrograma abrangendo proveniências de Itália e Sicília (também Córsega e Sardenha), o ramal S3-S4-S7 abrangendo Argélia e Tunísia, Sardenha e Córsega, e Provença, e o ramal S5-S6 abrangendo a Aquitânia, quase toda a Península Ibérica e Marrocos (S5 é provavelmente o que coincide com o “suber” predominante em Marrocos e na Península Ibérica [Belahbib et al. 2001, Jiménez et al. 2004], e seria altamente desejável que houvesse um esforço de uniformização das nomenclaturas). O ramal itálico, por sua vez, é fortemente aparentado com um existente em Q. cerris, o que é evidência eloquente da preponderância da partilha de refúgios glaciares na fixação de haplótipos comuns, muito mais credível do que a interpretação dos autores deste estudo, segundo a qual o sobreiro teria origem na Itália [Lumaret et al. 2005]. Finalmente, confirma-se a ausência de haplótipos “suber” nos sobreiros da costa oriental da Península Ibérica (figura 2.9) e na Catalunha francesa.
A introgressão de azinheira em sobreiro (e, em menor escala, no sentido inverso), com base nos estudos de DNA citoplásmico citados, parece incontestável. Pelas analogias com outros Quercus (cf. parte II secção D2) constituiria apenas mais um exemplo do que provavelmente é uma característica do género, não fossem dois aspectos que o colocam àparte: as contradições que tem com a classificação destas duas espécies em subgéneros diferentes, feita pela Flora Europæa [Tutin 1964], que como se viu na parte I é um esquema taxonómico filogeneticamente incorrecto mas reiteradamente faz perceber este caso de hibridismo como uma pretensa anomalia [Toumi & Lumaret 1998, Belahbib et al. 2001, Collada et al. 2002, Jiménez et al. 2004]; e as implicações de gestão e ordenamento, nomeadamente quanto ao futuro da azinheira no que hoje são grandes extensões de montado misto em Portugal.
Até agora conhece-se apenas a exploração preliminar de marcadores de nSSRs dinucleotídicas (GA)n [Soto et al. 2003], usando sequências conservadas entre Quercus como alvos dos primers (tabela 2.4). A análise de amostras de folhas dos descendentes obtidos por polinização controlada confirmou a codominância dos marcadores utilizados. Com uma amostragem por locus de apenas 18 cromossomas de sobreiro e 22 de azinheira, seria temerário dizer que qualquer um dos 6 loci propostos já estão validados como ferramentas de diagnóstico de hibridismo: o elevado polimorfismo dos fragmentos amplificados, que leva a considerar a hipótese de serem encontrados em amostragens maiores outros alelos eventualmente não-discriminantes, e o reduzido distanciamento entre “alelos” presumivelmente discriminantes (por causa das “bandas-fantasma” distanciadas da “real” de 1 par nucleotídico ou até mais [Hornero et al. 2001]) faz com que, apesar de promissora, esta abordagem ainda deva requerer mais loci para a detecção em larga escala de híbridos e principalmente introgressões. Este estudo merece atenção também pelo facto de introduzir nas duas espécies uma série de marcadores altamente polimórficos, de modo a permitir estudar a sua biologia de reprodução da mesma maneira que foi feito noutras espécies (cf. parte II secção D1).
Tabela 2.4 — Resumo dos polimorfismos de nSSRs de sobreiro e azinheira em 6 loci propostos por Soto et al. [2003] para uso na discriminação interspecífica entre sobreiro, azinheira, e seus híbridos.
|
Quercus suber (N = 9) / Quercus ilex (N = 11) |
|||||
|
MSQ13 |
MSQ4 |
QpZAG9 |
QpZAG15 |
QpZAG36 |
QpZAG46 |
nº total de alelos |
1/7 |
3/2 |
2/5 |
3/12 |
8/5 |
5/4 |
nº alelos discriminantes (e com diferença > 1 bp) |
1/7 (1/7) |
2/2 (2/1) |
2/5 (1/3) |
2/11 (2/11) |
7/4 (5/3) |
5/4 (4/3) |
freq. do alelo comum |
1,00/0,27 |
0,78/0,82 |
0,89/0,55 |
0,56/0,23 |
0,33/0,46 |
0,44/0,50 |
diagn. interespecíficoa |
+ |
– |
– |
– |
– |
– |
diagn. intra-específicob |
– / + |
+ / + |
+ / + |
+ / + |
+ / + |
+ / + |
a poucos alelos, todos discriminantes e com uma diferença entre espécies acima de 1 par nucleotídico (bp), é favorável à detecção de híbridos e introgressões (+)
b um elevado número de alelos, especialmente se o mais comum tiver frequência baixa, torna o locus propício para análise parental (+)
Como já discutido na parte II, secção D2, os híbridos de sobreiro e azinheira são os que melhor se prestaram até agora à análise genética, pela existência de marcadores monomórficos discriminantes e codominantes, podendo assim já contar-se com uma bateria de vários loci isoenzimáticos [Oliveira et al. 2003 e presente trabalho], a região central dos loci rDNA [Bellarosa et al. 2005] e, espera-se, um ou mais marcadores nSSR [Soto et al. 2003].