Parte I: Taxonomia do género Quercus
O género Quercus L., onde se incluem o sobreiro — Q. suber L. — e a azinheira — Q. ilex L. ssp.
rotundifolia (Lam.) O. Schwz. ex Tab. Morais
![]() — é classificado na família das Fagáceas
(Angiospérmicas). O recente esquema taxonómico elaborado pelo Angiosperm Phylogeny Group [APG
1998, Soltis et al. 2000] propõe a seguinte hierarquia de níveis taxonómicos intermédios (evidencia-se
em cada nível, com sublinhado, o grupo onde se incluem os Quercus):
— é classificado na família das Fagáceas
(Angiospérmicas). O recente esquema taxonómico elaborado pelo Angiosperm Phylogeny Group [APG
1998, Soltis et al. 2000] propõe a seguinte hierarquia de níveis taxonómicos intermédios (evidencia-se
em cada nível, com sublinhado, o grupo onde se incluem os Quercus):
Angiospermæ (clades supra-ordinais Monocotiledonæ, Eudicotiledonæ, ordens Ceratophyllales, Laurales, Piperales, Magnoliales, famílias Amborellaceæ, Nympheaceæ, Winteraceæ, etc.);
Eudicotiledonæ (clades supra-ordinais Asteridæ e Rosidæ, esta incluindo Eurosidæ I e II, ordens Geraniales, Caryophyllales, Santalales, Saxifragales, Proteales, Ranunculales, famílias Dilleniaceæ, Buxaceæ, etc.);
Eurosidæ I (ordens Cucurbitales, Fabales, Fagales, Malpighiales, Oxalidales, Rosales, família Celastraceæ, etc.);
Fagales Engl. (famílias Fagaceæ, Betulaceæ, Casuarinaceæ, Juglandaceæ, Myricaceæ, etc.);
Fagaceæ: árvores, raramente arbustos, monóicos, anemófilos ou mais raramente entomófilos; folhas simples, alternas, pecioladas, peninérvias, com estípulas caducas; flores masculinas formando amentos mais ou menos alongados, perianto sepalóide dividido em 4–6 lóbulos, (4)6–20(40) estames; flores femininas solitárias ou em grupos de 2 ou 3, rodeadas por um invólucro basal de que deriva a cúpula do fruto, perianto dividido em 4–6 lóbulos, ovário ínfero geralmente trilocular, com 2 rudimentos seminais por lóculo, 3 ou 6 estiletes (às vezes com 6 ou 9 lóculos e 6 a 9 estiletes); frutos em aquénio, de pericarpo coriáceo, solitários ou em grupos de 2 ou 3, rodeados por uma cúpula acrescente [Tutin 1964, Franco 1990];
Quercus L.: flores masculinas formando amentos longos e pendentes; inflorescências femininas posicionadas separadamente dos amentos; fruto contendo 1 aquénio oblongo ou elipsóide de germinação hipógea, cúpula escamosa abarcando somente a base do aquénio, formando como que um dedal; gemas ovóides [Tutin 1964, Galliano 1987, Franco 1990, Nixon 1993].
Outros géneros de fagáceas representadas na Europa e Bacia Mediterrânica são Fagus L. (faias) e Castanea Mill. (castanheiros).
1) Subdivisões do género Quercus
A taxonomia infragenérica dos Quercus tem relevância para o estudo dos híbridos entre sobreiro e azinheira sob dois pontos de vista: por um lado, porque os caracteres diagnosticantes de cada grupo subgenérico são úteis na compilação de caracteres diagnosticantes entre as duas espécies e os seus híbridos, a qual será tratada na secção C1 da III parte; por outro, pela relação que tem o nível hierárquico da separação entre as duas espécies e a plausibilidade da ocorrência de híbridos, que no esquema taxonómico adoptado pela Flora Europæa [Tutin 1964] não “devem” ocorrer.
A literatura europeia referencia-se geralmente ao esquema adoptado pela Flora Europæa [Tutin 1964], proposto por O. Schwarz [1936]. Nele se consideram 3 subgéneros autóctones para a Europa e 1 exótico (Erythrobalanus) (figura 2.1a), mas está longe de considerar o género no seu todo, e a literatura norte-americana ignora-o. Como comparação, no esquema proposto por Krüssmann com base no sistema de K. Prantl para toda a espécie [Krüssmann 1978, Kleinschmit 1993], as espécies europeias pertencem a um só subgénero, Lepidobalanus (figura 2.1b).
 Figura 2.1 — Exemplos da subdivisão do género Quercus em categorias supra-específicas, ilustrando a classificação nessas categorias das espécies ibéricas (autóctones
excepto no caso de Q. rubra L. e Q. cerris L.). a) Esquema adoptado pela Flora Europæa.
b) Esquema proposto por Krüssmann.
Figura 2.1 — Exemplos da subdivisão do género Quercus em categorias supra-específicas, ilustrando a classificação nessas categorias das espécies ibéricas (autóctones
excepto no caso de Q. rubra L. e Q. cerris L.). a) Esquema adoptado pela Flora Europæa.
b) Esquema proposto por Krüssmann.Mais recentemente, as análises cladísticas de Nixon contemplaram apenas 2 subgéneros, Cyclobalanopsis e Quercus, neste separando-se três secções (Quercus = Lepidobalanus, onde os grupos Ilex e Cerris-Suber poderiam formar uma subsecção àparte, Lobatæ = Erythrobalanus e Protobalanus só de espécies americanas) [Nixon 1993, Borgardt & Pigg 1999], mas com base na variação molecular [Manos et al., 1999, 2001] esse esquema também não será o mais adequado, pois os subgéneros Cerris e Schlerophyllodrys da Flora Europæa constituiriam uma ramificação provavelmente tão precoce como os Cyclobalanopsis (figura 2.2, Apêndice II secção A).
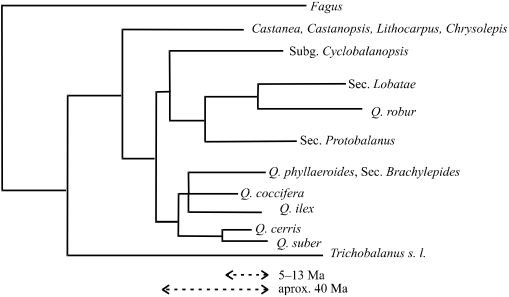 Figura 2.2 — Esquema da filogenia das Fagáceas baseada na análise das sequências
das regiões ITS de rDNA, simplificado a partir de Manos et al. [2001]. Usando como
referência Q. suber, ilustram-se 2 critérios de calibração do tempo geológico para estes
dados (Ma, milhões de anos).
Figura 2.2 — Esquema da filogenia das Fagáceas baseada na análise das sequências
das regiões ITS de rDNA, simplificado a partir de Manos et al. [2001]. Usando como
referência Q. suber, ilustram-se 2 critérios de calibração do tempo geológico para estes
dados (Ma, milhões de anos).Segundo os resultados de Manos et al. [2001] com as sequências dos ITS (internal transcribed spacers) do rDNA, a divergência entre sobreiro e azinheira é cerca de 50% superior à que os separa dos táxones mais aparentados com cada um (Q.cerris e Q. coccifera, respectivamente), aproximadamente a mesma que separa Q. robur (carvalho roble) de Q. alba L., e menos de 60% da que separa o sobreiro do roble. Assim, em termos filogenéticos, o sobreiro não será um parente muito afastado da azinheira dentro do género Quercus, mas também não se conta entre os exemplos de maior proximidade. Por outro lado, o estudo da mesma região cromossómica num pequeno conjunto de espécies europeias, utilizando modelação da estrutura secundária dos transcritos (primariamente para aperfeiçoar o alinhamento das sequências) reafirma a existência de 3 grupos conforme a classificação da Flora Europæa [Bellarosa et al. 2005], mas sem ser contraditória com o esquema geral de Manos et al. [2001], que tem a vantagem de incluir um número muito considerável de espécies e assim permitir uma melhor avaliação dos parentescos dentro do género.
Apesar de ser necessária muita precaução na interpretação das filogenias baseadas em caracteres moleculares, elas têm o potencial de virem a dissipar a arbitrariedade inerente aos esquemas clássicos, deste modo podendo avaliar-se melhor qual o grau de afastamento filogenético entre sobreiro e azinheira, o que acarreta a priori duas considerações pertinentes para o presente estudo:
– será tanto mais fácil encontrar caracteres moleculares discriminantes entre as duas espécies quanto mais prolongada tiver sido a oportunidade para uma divergência a nível nucleotídico e, consequentemente, a nível proteico; e
– num maior afastamento filogenético, haverá menores probabilidades de ocorrência de híbridos pela acumulação de barreiras genéticas, do que entre espécies mais próximas entre si.
O registo fóssil [Borgardt & Pigg 1999, Kashani & Dodd 2002, Bellarosa 2003] faz associar a diversificação dos Quercus à transição climática de Eocénico para o Oligocénico (há cerca de 40 milhões de anos), enquanto formas muito semelhantes às espécies actuais ocorreriam já durante o Miocénico (há 5 a 13 × 106 anos, figura 2.2). Tendo em conta a alternância climática com fases glaciares, e consequentes recuos da flora das regiões temperadas para refúgios localizados a Sul já no Plistocénico (a partir de há 1,5 × 106 anos), é pouco provável que se possa deduzir a origem geográfica das espécies de Quercus que não sejam extremamente recentes.
2) As categorias taxonómicas e o hibridismo
A noção de que espécies diferentes não se cruzam naturalmente — ou antes que a reprodução sexuada se realiza normalmente apenas entre indivíduos da mesma espécie — herdada dos tempos pré-mendelianos [Darwin 1872] e formulada em termos genéticos por Dobzhansky e sobretudo por Mayr para integrar o conceito de espécie na teoria neodarwinista [Van Valen 1976, Dobzhansky et al. 1977], encontra no género Quercus um dos contra-exemplos mais notáveis [Burger 1975]. O hibridismo entre Quercus é de tal maneira conspícuo, que os imperativos genéticos/reprodutores da definição neodarwinista se tornam incompatíveis com a delimitação de espécies essencialmente morfológica e ecológica, a qual permanece como o critério mais viável para uma taxonomia com sentido prático e consistência filética: nos Quercus, a definição neodarwinista tenderia a corresponder aos subgéneros (ou às secções, segundo os esquemas taxonómicos), e não ao que a nomenclatura botânica considera espécies [Burger 1975]. Por vezes preconiza-se considerar espécies que hibridam como sendo subespécies [Kleinschmit & Kleinschmit 2000], mas isso cria mais problemas do que pretende resolver (se é que resolve), nomeadamente por subverter (pelo menos no caso do par em questão, Q. robur e Q. petræa) alguns dos pressupostos consensuais para a definição de subespécie [Lawrence 1951]. E o termo semi-espécie proposto para o caso particular dos muitos Quercus que hibridam [Burger 1975] também parece inadequado, por ser-lhe implícita uma imaturidade do processo de especiação que é contradita pelos factos [Van Valen 1976]. Sem negar a utilidade do conceito neodarwinista baseado em critérios ligados à reprodução, a lição que parece tirar-se das dificuldades da sua aplicação nos Quercus é a necessidade de aplicar os conceitos com flexibilidade [Burger 1975].
Uma definição de espécie que procura conferir-lhe a maior flexibilidade e ao mesmo tempo circunscrevê-la em critérios precisos e verificáveis é a de Van Valen [1976], para a qual o critério de delimitação das diferentes espécies é o de cada uma ocupar uma zona adaptativa que é única dentro da sua área de distribuição. Este conceito de espécie, dito ecológico (mas também filogenético), não impõe qualquer restrição ao fluxo genético entre espécies simpátricas no seu estado natural, desde que se mantenham como linhagens evolutivas separadas, isto é, ocupando zonas adaptativas diferentes.
Um sistema alternativo de categorização de formas aparentadas, sem referência explícita a espécies mas resultando de estudos de hibridação controlada, são as chamadas categorias Bio-Sistemáticas definidas por Turresson [Lawrence 1951]. Às “espécies” que formam híbridos férteis e viáveis atribui-se a categoria de ecospécies, e ao conjunto entre ecospécies que hibridam a categoria de cenospécie. Uma cenospécie corresponderia a priori, aproximadamente, ao nível de secção ou de subsecção. Na nomenclatura de Van Valen prefere-se o termo multi-espécie, pelo facto da cenospécie se definir fora do estado natural [Van Valen 1976].
Não é no sentido da flexibilidade preconizada que vai a noção, mais ou menos explícita em diversos textos [Toumi & Lumaret 1998, Belahbib et al. 2001, Collada et al. 2002], que os Quercus hibridam frequentemente apenas dentro de secções do mesmo subgénero. Cabe então perguntar qual o esquema taxonómico dentro do qual iria dar-se crédito a tal afirmação. Os híbridos que se formam entre sobreiro e azinheira são um exemplo notável da dificuldade: pelo esquema de Schwarz adoptado pela Flora Europæa, trata-se de híbridos entre membros de diferentes subgéneros, e como são em geral viáveis e férteis, esta contradição levou o próprio Schwarz, segundo relata Natividade [1947], a negar a sua existência!
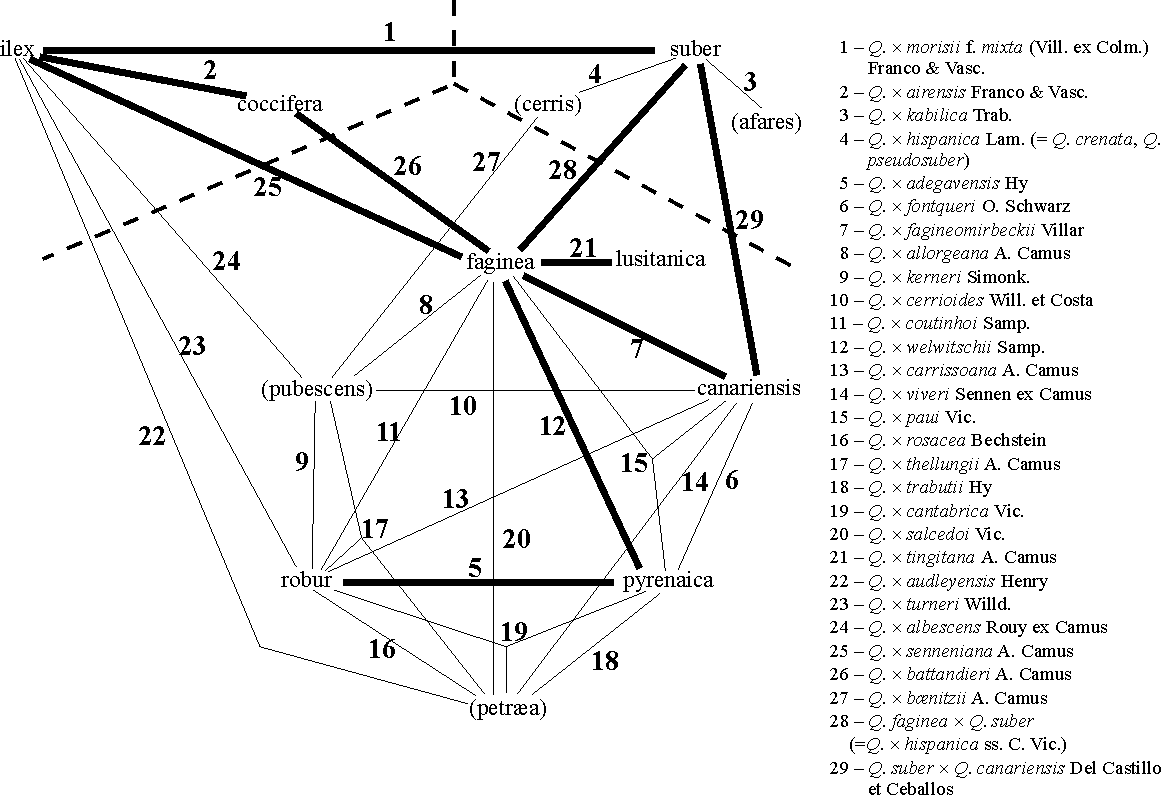 Figura 2.3 — Compilação de híbridos registados, entre espécies de Quercus que ocorrem na
Península Ibérica [Coutinho 1888, Natividade 1950, Vasconcelos & Franco 1954, Galliano 1987,
Franco 1990, Hélardot s.d.]. Apenas se indicam os restritivos específicos, ficando entre parêntesis os
das espécies que não ocorrem em Portugal (além disso, Q. afares Pomel não é ibérica). A traço mais
grosso indicam-se as combinações que podem a priori encontrar-se em Portugal. A tracejado, limites
subgenéricos segundo o esquema de Schwarz [Tutin 1964]. Todas as designações envolvendo Q. ilex
referem-se em princípio à subespécie rotundifolia. 1: híbridos de sobreiro e azinheira; 2: híbridos
dentro do subgénero Sclerophyllodrys; 3–4: híbridos dentro do subgénero Cerris; 5–21: híbridos
dentro do subgénero Quercus; 22–26: híbridos entre os subgéneros Sclerophyllodrys e Quercus;
27–29: híbridos entre os subgéneros Cerris e Quercus.
Figura 2.3 — Compilação de híbridos registados, entre espécies de Quercus que ocorrem na
Península Ibérica [Coutinho 1888, Natividade 1950, Vasconcelos & Franco 1954, Galliano 1987,
Franco 1990, Hélardot s.d.]. Apenas se indicam os restritivos específicos, ficando entre parêntesis os
das espécies que não ocorrem em Portugal (além disso, Q. afares Pomel não é ibérica). A traço mais
grosso indicam-se as combinações que podem a priori encontrar-se em Portugal. A tracejado, limites
subgenéricos segundo o esquema de Schwarz [Tutin 1964]. Todas as designações envolvendo Q. ilex
referem-se em princípio à subespécie rotundifolia. 1: híbridos de sobreiro e azinheira; 2: híbridos
dentro do subgénero Sclerophyllodrys; 3–4: híbridos dentro do subgénero Cerris; 5–21: híbridos
dentro do subgénero Quercus; 22–26: híbridos entre os subgéneros Sclerophyllodrys e Quercus;
27–29: híbridos entre os subgéneros Cerris e Quercus.Para exemplificar a complexidade do hibridismo entre os Quercus, ilustra-se na figura 2.3 uma compilação dos híbridos espontâneos entre espécies ibéricas, muitos deles férteis, senão todos. Registam-se, por exemplo, só entre 6 espécies do subgénero Quercus s.s. (esquema da Flora Europæa, excluindo Q. lusitanica), 13 combinações duas a duas em 15 possíveis, às quais se vêm acrescentar mais 9 envolvendo outras espécies ibéricas. Como se pode ver nessa figura, os híbridos de sobreiro e azinheira são apenas uma de várias combinações entre espécies de diferentes subgéneros de acordo com o esquema de Schwarz. Conclui-se assim que a referência ao esquema taxonómico da Flora Europæa, para além de ser contraditória com esquemas mais actualizados e “objectivos”, dificulta uma correcta apreciação dos fenómenos de hibridismo neste género.
Sendo as possibilidades de hibridismo entre Quercus tão diversificadas, o facto das espécies manterem-se diferenciadas em diversos aspectos (morfológicos, ecológicos, geográficos) leva a postular-se o prevalecimento de selecção disruptiva tendente à eliminação de formas intermédias descendentes de híbridos [Nason et al. 1992, González-Rodríguez & Oyama 2005], podendo ou não ser potenciada por uma menor fertilidade desses híbridos [Nason et al. 1992, Jiggins & Mallet 2000]. É provável que muitas das espécies que actualmente hibridam já fossem entidades separadas no Pliocénico tardio mas tenham estado sujeitas a sucessivos movimentos de expansão/recolonização e recuo/fragmentação no decurso dos ciclos de glaciação-interglaciação que caracterizaram o Plistocénico, dando a oportunidade repetida para contactos e hibridação [González-Rodríguez et al. 2004b]. Neste cenário admite-se, por exemplo, o postulado de sistemas de espécies coevoluindo em habitats partilhados, e onde o fluxo genético entre elas poderia constituir um factor de coesão, nomeadamente pela introgressão de genes relevantes adaptativamente, alargada sobre as várias populações duma mesma multi-espécie [Burger 1975]. Tais sistemas tendem a perpetuar-se nesses habitats e beneficiam da complementaridade ecológica das diferentes espécies, e em ambientes extremos, por exemplo refúgios microclimáticos durante os períodos de glaciação, essa coesão poderia ainda ser mais reforçada. A verificar-se repetidamente a existência de tais sistemas nos Quercus, nem mesmo a definição proposta por Van Valen [1976] é suficientemente flexível para as espécies deste género, e pode ser muito difícil definirem-se marcadores genéticos discriminantes entre membros duma mesma multi-espécie. Uma consequência desta situação, é que a diferenciação genética entre os membros destas multi-espécies pode limitar-se a apenas uma componente muito limitada do genoma que responde à selecção disruptiva, de maneira que nem sempre é de esperar que a diferenciação morfológica seja acompanhada pelos marcadores genéticos dispersos no genoma [González-Rodríguez et al. 2004a].
Por outro lado, em certas condições pode surgir uma nova espécie a partir de uma linhagem híbrida, por um processo de segregação de novos recombinantes entre as espécies parentais chamado “especiação híbrida homoplóide”. Identificaram-se alguns parâmetros fundamentais para o sucesso deste processo, designadamente: fertilidade do híbrido inicial, percentagem de autopolinização no híbrido e sua descendência, vantagem selectiva dos genótipos recombinantes, semelhança cariotípica entre as espécies parentais, e extensão geográfica do contacto entre estas [Rieseberg 1997]. Nos Quercus em geral é conhecida a fertilidade de muitos híbridos (ao ponto de constituirem-se grexes de pelo menos 3 espécies, caso dos táxones 15, 17 e 19 da figura 2.3), assim como a aparente uniformidade cariotípica (2n = 2x = 24), pelo menos dentro das espécies europeias [Tutin 1964, Franco 1990]; mas no que respeita à taxa de autopolinização nos híbridos, a supor-se que acompanha a tendência do género para preferir a polinização cruzada, ela será baixa, isso não é obstáculo desde que o factor selectivo seja suficientemente forte [Rieseberg 1997].
Este modelo de especiação actuaria assim num sentido oposto ao da selecção disruptiva, produzindo zonas híbridas unimodais [Jiggins & Mallet 2000] onde conseguiriam sobreviver os híbridos (e talvez parte das linhagens deles derivadas), especialmente se isoladas em relação às espécies deles progenitoras; pode assim considerar-se a aparente facilidade de hibridação neste género como um factor de favorecimento da especiação por hibridismo, isto é, a formação de novas espécies estabilizadas a partir de híbridos (hibri-espécies), desde que para a ocupação de novas zonas adaptativas, por exemplo em habitats extremos em relação a pelo menos uma das espécies parentais ou em locais de perturbação ecológica, incluindo antropogénica [Nason et al. 1992, Rieseberg 1997, Huxel 1999, Valbuena-Carabaña et al. 2005]. Os processos de especiação envolvem a recursividade entre a divergência genética e os seus efeitos no isolamento reprodutor e consequente redução do fluxo genético, num fenómeno de feedback positivo [Rice & Hostert, 1993], e a rápida segregação de novas combinações genotípicas a partir de híbridos férteis serviria de base para o processo. Pode imaginar-se que no género Quercus, o mais diversificado das Fagales com as suas mais de 500 espécies [Goværts & Frodin 1998], haja um historial (e potencial para futuro) de especiação a partir de híbridos, e são frequentes as referências botânicas à possível origem híbrida dalgumas espécies. Um exemplo recentemente relatado no género Pinus oferece detalhes geográficos, genéticos e ecológicos que ilustram muito bem o modelo de especiação híbrida homoplóide [Wang et al., 2001], e pelo menos um estudo genético invoca o mesmo processo em Quercus [Mir et al. 2006] (cf. parte II secção D2).