Feita a descrição dos marcadores isoenzimáticos desenvolvidos, em termos de variação dos zimogramas, limitações de revelação e também, nalguns casos, da respectiva hereditariedade, torna-se possível aplicá-los à investigação de diversas questões, com as amostragens feitas.
Como referido na secção anterior, o recurso a hipóteses genéticas plausíveis sobre a hereditariedade dos marcadores polimórficos, seja a nível interespecífico como intra-específico, permite identificar indivíduos cuja etiquetagem é errada. As causas possíveis para a ocorrência de indivíduos erroneamente etiquetados, dentro do esquema de amostragem adoptado, são essencialmente três:
i) intrusão de sementes de origem diversa antes da sua colheita, deslocadas após a sua queda ao solo por acidentes do terreno, acção de animais ou outros agentes (contaminantes in situ);
ii) erros de etiquetagem no processo de sementeira, no período de crescimento em viveiro, ou no processamento para análise isoenzimática (contaminantes ex situ);
iii) erros no registo dos zimogramas.
A utilização de fotografias legendadas permitiu corrigir a posteriori alguns casos de erro do terceiro tipo, e o registo da morfologia das plantas no acto de colheita de amostras de folhas permitia prever em que casos se encontraria um zimograma contraditório com a espécie implícita na etiqueta. De resto, só com os zimogramas se pôde ter alguma noção sobre a extensão das contaminações, a qual, como já referido a propósito dos resultados da tabela 4.10, pelas limitações inerentes aos marcadores enzimáticos utilizados, ainda assim tem de entender-se por defeito. A tabela 4.11 resume a evidência obtida para os contaminantes in situ.
Tabela 4.11 — Incidência de contaminantes in situ durante o presente estudo. Entre parêntesis, dimensão da amostra para cada caso. Os contaminantes interspecíficos referem-se à presença de plantas duma espécie dispersas entre a amostragem doutra espécie, e os marcadores intra-específicos (PGI em sobreiro e PGM em azinheira) à detecção de contaminantes pela incongruência entre os genótipos das plantas jovens e os das árvores às quais era atribuída a sua origem materna.
|
Mitra |
Feijoas |
Alfaiates |
Global |
||||
Contaminantes interspecíficos |
0,0% |
(351) |
1,1% |
(282) |
1,4% |
(554) |
0,9% |
(1187) |
Marcadores PGI (sobreiro) |
1,9% |
(53) |
2,2% |
(46) |
3,8% |
(160) |
3,1% |
(259) |
Marcadores PGM (azinheira) |
8,1% |
(223) |
4,4% |
(90) |
19,3% |
(181) |
11,5% |
(494) |
Conforme sugerido na secção anterior, é possível que a estimativa de contaminantes entre azinheiras no povoamento de Alfaiates esteja inflacionada por um erro de classificação da árvore 21, onde ocorreram cerca de metade do total de contaminantes desta espécie. Admitindo a hipótese desta azinheira ser ac, a percentagem em Alfaiates baixa para 4,4% e a global para 6,1%.
A detecção de contaminantes in situ interspecíficos contrasta pela prevalência muito mais baixa; e a distribuição desses 11 contaminantes também foi heterogénea: 8 eram azinheiras dispersas na amostragem da árvore 20 do povoamento de Alfaiates (sobreiro), aliás vizinha da 21 (azinheira), ambas situadas no sopé da encosta onde assenta este povoamento (cf. “Materiais e Métodos” parte II secção A1, e Apêndice I secções A1 e B3); as outras 3 eram de Feijoas, 2 sobreiros e 1 azinheira, em 3 famílias diferentes.
Em termos de contaminantes ex situ, houve 4 caixas que eram compostas por sobreiros, mas cuja etiquetagem se referia a uma azinheira. Todas foram recebidas com a derradeira remessa de plantas, com sinais de ter havido colocação de novas etiquetas identificadoras, o que é sugestivo de ser uma troca ocorrida já após as sementeiras, em fase tardia da sua manutenção no viveiro.
Também num dos povoamentos de referência se registou 1 planta contaminante de azinheira (amostra 20Q) no de Santiago do Cacém.
Entre os adultos observados, os únicos híbridos foram aqueles que já estavam referenciados como tal antes de serem analisados (tabela 4.12):
Tabela 4.12 — Fenótipos isoenzimáticos dos híbridos analisados no presente estudo.
Árvore |
Amostras |
DIA |
MDH |
GSR |
PGM |
PGD |
PGI |
EST |
Comentários |
ALC |
X24 |
X |
X(X) |
X |
|
12 |
Az+b |
12 |
extracção 41 |
CAB |
X25 |
X |
X(X) |
X |
|
|
+ |
2 |
extracção 41 |
MRM |
X10, X27 |
X |
X(X) |
X |
|
12 |
Az |
12 |
amostra X10 deu Az em DIA; extracções 28 e 42 |
SES |
X4, X8, X13, X20 |
X |
X(X) |
X |
|
12 |
Az |
12 |
extracções 0, 7, 28 e 39 |
SM1 |
X2, X15, X22 |
X |
X(X) |
X |
(ad) |
12+b |
+ |
2 |
amostra X15 deu 12 em EST; fenótipo PGM deduzido a partir da descendência (tabela 4.10); extracções 0, 31, 39 |
SM2 |
X3, X16, X23 |
X |
X(X) |
X |
d |
12 |
Az |
2 |
extracções 0, 31, 39 |
SM3 |
X5, X17 |
X |
X(X) |
X |
cd |
12+b |
Az |
2 |
extracções 7, 31 |
SM4 |
X14, X21 |
X* |
X(X) |
X |
|
12 |
Az |
12 |
extracções 28 e 39 |
VR |
X26 |
X? |
X(X) |
X |
|
12 |
Az+b |
2 |
extracção 41 |
Em todos se verificaram zimogramas DIA, MDH (ânodo e cátodo) e GsR híbridos. Já a análise para a actividade PGM trouxe 2 problemas: apesar da repetida amostragem em vários casos, só foi observada directamente em 2 híbridos (e indirectamente num terceiro, graças às distribuições na descendência); e na árvore SM2 o fenótipo é idêntico ao dos sobreiros, o que foi confirmado pela observação da respectiva descendência (tabela 4.10). Pode assim tratar-se dum descendente de híbrido, a exemplo dos que foram observados no presente estudo (cf. parte D deste capítulo), ou então, se for um verdadeiro híbrido, o gâmeta de azinheira que lhe deu origem teria sido portador do alelo raro Pgm5. Finalmente, pode verificar-se que os marcadores menos específicos (PGD, PGI e EST), em conjunto, permitiriam distinguir 8 destes híbridos dos sobreiros (a excepção foi CAB).
Tanto em folhas como em sementes, nenhum híbrido foi observado. A única conclusão que se pode tirar, qualquer que seja o conjunto de amostras que se considere, é que a probabilidade de incidência de híbridos é majorada pelo valor P, tal que NP < –loge(1 – α), em que N é o número de indivíduos analisados e α a probabilidade de obter pelo menos 1 híbrido (distribuição de Poisson). Na tabela 4.13 listam-se os valores estimados, considerando apenas os conjuntos onde se fez análise de folhas, visto que a amostragem de sementes foi muito limitada.
As estimativas de P, muito baixas para cada espécie em cada povoamento misto em função dos valores de N deste estudo, podem ser majoradas por um valor tão baixo quanto 0,004% (α = 5%), considerando uniformidade entre povoamentos e entre espécies. Mesmo relaxando o nível de erro α para 10% ou 20%, a estimativa permanece bastante baixa: como exemplo, e admitindo que as azinheiras são as mais prováveis receptoras na hibridação, o majorante de P situa-se em 0,043% com α = 20% (o que se presume dar uma margem bastante confortável em relação ao valor real).
Tabela 4.13 — Cálculo do majorante da probabilidade P de incidência de híbridos nos povoamentos mistos estudados, para níveis de erro α de 5%, 10% e 20%, com base nas observações em plantas germinadas de sementes dos três povoamentos. Excluem-se apenas as que pertencem a caixas cuja etiquetagem era errada (contaminantes ex situ).
|
|
|
|
P |
||
Espécie |
Frutificação |
Local |
N |
α = 5% |
α = 10% |
α = 20% |
Azinheiras |
1997 |
Mitra |
75 |
<0,068% |
<0,140% |
<0,298% |
1998 |
Mitra |
223 |
<0,023% |
<0,047% |
<0,100% |
|
Feijoas |
91 |
<0,056% |
<0,116% |
<0,245% |
||
Alfaiates |
211 |
<0,024% |
<0,050% |
<0,106% |
||
Global |
525 |
<0,010% |
<0,020% |
<0,043% |
||
Sobreiros |
1998 |
Mitra |
116 |
<0,044% |
<0,091% |
<0,192% |
Feijoas |
179 |
<0,029% |
<0,059% |
<0,125% |
||
Alfaiates |
343 |
<0,015% |
<0,031% |
<0,065% |
||
Global |
638 |
<0,008% |
<0,017% |
<0,035% |
||
Ambas |
1998 |
Mitra |
339 |
<0,015% |
<0,031% |
<0,066% |
Feijoas |
278 |
<0,018% |
<0,038% |
<0,080% |
||
Alfaiates |
554 |
<0,009% |
<0,019% |
<0,040% |
||
Global |
1163 |
<0,004% |
<0,009% |
<0,019% |
||
Considerando como modelos de referência as espécies “puras” e os híbridos, os casos que apresentem combinações de fenótipos enzimáticos dos diferentes modelos foram tratados a priori como exemplos possíveis de introgressão.
1) Povoamentos mistos e de referência
Houve 6 amostras de árvores adultas consideradas ambíguas pelo conjunto de resultados obtido, 3 das quais (amostras 1L, 2G e 2N) devido a erros de avaliação dos zimogramas EST e MDH, numa fase muito precoce do trabalho, e que foram esclarecidos em novas amostragens. As outras referem-se a 3 azinheiras, a árvore 30 do povoamento de Feijoas, cuja PGM era bd, e as árvores 11 e 36 do povoamento de Alfaiates, que são DIA “X”, e das quais parte da descendência era do mesmo fenótipo — isto é, como os híbridos em DIA mas típico das azinheiras em todos os outros marcadores analisados. Em ambas as situações poderá persistir a dúvida entre tratar-se de introgressão dum gene de sobreiro e a ocorrência dum alelo raro de azinheira. No caso da banda d de PGM, a sua ocorrência nas descendências de Testa, Azeiteiros e Alfaiates (cf. secção E) é consistente com a existência dum alelo raro de azinheira que codifica uma actividade de comigra com a de sobreiro. Alternativamente, considerar-se-ia uma introgressão do alelo de sobreiro, e no caso de Alfaiates (dado que a banda d não foi observada em nenhum dos adultos) transmitido por via polínica.
A evidência de aparente introgressão do gene para DIA de sobreiro pode ser avaliada nas linhagens das árvores 11 e 36 de Alfaiates, sob o ponto de vista da hipótese genética enunciada na secção B1, em que se prevê que metade dos descendentes seja portador do gene de sobreiro e assim apresentem o zimograma “X”. Na tabela 4.14 resume-se a informação disponível e o resultado do teste estatístico correspondente.
Tabela 4.14 — Expressão de DIA como zimograma “X” nas descendências das árvores 11 e 36 de Alfaiates. A coluna de probabilidade refere-se à estatística χ2 (1 g.l.) com hipótese nula de 50% de descendentes expressando esse zimograma. O teste de homogeneidade entre as descendências deu χ2 = 0,64 (P = 0,42, 1 g.l.).
Descendência |
Total |
Observados |
Esperados |
χ2 |
P |
A11 |
14 |
6 |
7,0 |
0,07 |
0,79 |
A36 |
37 |
10 |
18,5 |
6,92 |
0,009 |
A11+A36 |
51 |
16 |
25,5 |
6,35 |
0,012 |
Conclui-se que pelo menos em A36 há um desvio significativo entre as ocorrências observadas e os valores esperados pela hipótese genética enunciada na secção B1, o que não a invalida forçosamente, pois pode dever-se a selecção negativa sobre as plantas com DIA “X” (pode também dever-se à existência de contaminantes não detectados, apesar da expressão da PGM não evidenciar nenhum nestas descendências, cf. tabela 4.10).
Houve ainda 8 plantas provenientes dos povoamentos mistos com zimogramas ambíguos, das quais há a salientar 1 caso (amostra 37az pertencente à amostragem de 10 elementos atribuídos à azinheira 9 de Feijoas) onde a PGI foi classificada como “Az+b”, enquanto os marcadores DIA e MDH (anódico) eram “Az”. Excluindo a possibilidade de tratar-se dum contaminante doutra família, e visto que nenhum outro membro desta família apresenta esta banda PGI lenta, deduz-se que é um alelo transmitido por via paterna; e nessa, pode considerar-se a hipótese de introgressão (pois aparenta ser idêntica à banda b de sobreiro) ou duma actividade de azinheiras mais lenta (cf. tabela 4.8, figura 4.7 b).
Mais difíceis de explicar são 2 amostras (38c e 38i) atribuídas à árvore 19 da Mitra, cujo zimograma PGM é d. A árvore M19 tem zimograma PGM “ac” e por isso podem tratar-se de contaminantes intra-específicos, mas em todo o caso o restante dessas duas amostras é típico de azinheira. Houve ainda 2 amostras de sobreiros contíguas no mesmo gel (33Y e 33Z) que deram um zimograma PGI “a+b”, isto é, com as bandas a e b mas sem a banda intermédia característica dos heterozigóticos, tratando-se provavelmente de contaminação mútua, durante a colocação destas amostras no gel, entre uma de zimograma a e outra de zimograma b. Os restantes casos em povoamentos mistos prendem-se com a detecção da banda 1 de EST em sobreiros, mas pode tratar-se de erros de visualização numa fase relativamente precoce do trabalho.
Os restantes casos de ambiguidade de identificação provieram quase todos do povoamento de referência (sobreiros) de Santiago do Cacém, 20 no total, dos quais 17 dizem respeito à detecção da banda 1 em EST, todos numa fase relativamente precoce do trabalho e que talvez se possam atribuir a erros de visualização. Mais complexos, mas instrutivos, são os outros 3 casos, e ainda um (amostra 18r) do povoamento de referência Azeiteiros (tabela 4.15).
Tabela 4.15 — Fenótipos isoenzimáticos de 4 plantas consideradas de identidade ambígua, provenientes de povoamentos de referência. Na coluna PER, os fenótipos são banda 1 (presença ou ausência) / região 2 (variantes 0 e *, cf. figura 4.11). No texto refere-se ainda a actividade ACP-F (sistema 9) da amostra 20K.
Amostra |
DIA |
MDH |
PER |
PGI |
EST |
GSR |
PGM |
PGD |
15U |
X |
Sb(Az) |
+/0 |
Az |
12 |
Sb |
ac |
12 |
18r |
Az |
X? |
–/0 |
+ |
123 |
Az |
ac |
12 |
20K |
Az |
Sb(Sb) |
–/0 |
b |
2 |
Sb |
d |
12 |
30AB |
X |
Sb(Sb) |
–/0* |
b |
2 |
Sb |
d |
|
A amostra 15U, que tinha aspecto de azinheira, apresentou uma mistura de caracteres de sobreiro (MDH anódica, GsR), de azinheira (MDH catódica, PGI, EST e PGM) e de híbrido (DIA) que sugerem tratar-se dum caso de introgressão. Infelizmente, não foi possível reamostrar esta planta porque morreu depois de ser devolvida ao viveiro.
A actividade MDH anódica da amostra 18r foi considerada híbrida antes de ser conhecido o padrão “X” através dos híbridos, mas ao contrário destes apresentou-se difusa — e a ausência de banda 1 em PER contribuiu na altura para classificar esta amostra como ambígua; é possível, por isso, que essa avaliação fosse incorrecta e não haja introgressão neste caso.
As amostras 20K e 30AB, finalmente, apenas diferem do padrão de sobreiro pelo zimograma DIA, e, enquanto 30AB se assemelha à situação de introgressão proposta para as famílias 11 e 36 de Alfaiates (mas desta vez em sobreiro), 20K chega a apresentar o zimograma “Az” (e a actividade ACP-F, no sistema 9, migrou para uma posição intermédia entre sobreiros e azinheiras no mesmo gel). Situação análoga à da 20K, no que respeita ao zimograma DIA, foi observada em sobreiros preguentos da Herdade de Montinho, como se detalha a seguir.
Dos 3 povoamentos onde foi feita uma amostragem de sobreiros de cortiça preguenta, apenas no da Herdade de Montinho se detectaram zimogramas que podem ser indicadores de introgressão de azinheira. Essa evidência resume-se essencialmente ao marcador DIA, que em cinco árvores era “X” (árvores 2, 11, 12, 13, 19) e em duas era “Az” (números 15 e 18); uma segunda amostra de folhas foi colhida das árvores 11, 12 e 13, e confirmou esta observação. Todas estas árvores (excepto a número 2) são vizinhas entre si, ocupando, com outras aparentemente normais (números 14, 16, 17, 20 e 21, além de mais 4 próximas da número 18), uma área que não excede a de um quadrado com 50 metros de lado (cf. Apêndice I secção B4); este padrão de distribuição, e se se tratar de regeneração natural, sugere que estas árvores são aparentadas entre si por via materna.
Outro facto a assinalar nos sobreiros destes 3 povoamentos é a elevada proporção (22 a 27, num total de 45 amostras) dos que apresentavam a banda PER 1, o que é atípico da espécie (cf. tabela 4.5).
3) Desempenho dos marcadores isoenzimáticos nas análises de rotina
A tabela 4.16 resume o desempenho de cada uma das actividades analisadas em larga escala, de modo a permitir compará-las entre si em termos de consistência, especificidade e sensibilidade.
Tabela 4.16 — Percentagem de análises identificativas (consistência), especificidade e sensibilidade de cada um dos marcadores isoenzimáticos utilizados em larga escala no presente estudo (excluindo apenas as amostragens selectivas de sobreiros preguentos), referidas à possibilidade de detecção de hibridismo em sobreiros. Consistência abrange todas as análises, especificidade e sensibilidade apenas se refere à perspectiva de análise em sobreiros.
Sistema |
Marcador |
Número de anostras |
Consistência |
Especificidade |
Sensibilidade |
C |
DIA |
2242 |
96,2% |
100% |
100% |
C |
MDH |
2290 |
98,2% |
100% |
100% |
C |
PER |
2199 |
100,0% |
83,9% |
98,8% |
H |
GsR |
1591 |
70,6% |
100% |
100% |
H |
PGM |
1861 |
84,3% |
100% |
99,7% |
H |
PGD |
1934 |
85,8% |
100% |
12,2% |
R |
PGI |
2146 |
93,2% |
100% |
80,6% |
R |
EST |
2138 |
90,5% |
100% |
92,4% |
Os valores em PER referem-se apenas à banda 1. A quebra de sensibilidade em PGM deve-se à
ocorrência de 2 azinheiras (amostras 38c e 38i) com zimograma d. A sensibilidade em PGD refere-se às
azinheiras com banda b, em EST às que apresentaram banda 1 (que baixa para 48,6% nas descendências
de híbridos, mas a expectativa seria 26,0%, χ2 = 36,0, 1 g.l., P = 2 ×10–9). Quanto à sensibilidade em PGI,
refere-se a todas as azinheiras (plantas e sementes) que não eram “+”, mas há desvios importantes em
relação a este valor global: nos povoamentos mistos esta sensibilidade foi de 84,4% em Alfaiates, 94,3%
em Feijoas e 84,9% na Mitra; nos de referência foi mais baixo: 76,4% em Azeiteiros e 76,9% em Testa;
e nas descendências de híbridos, como seria de esperar
![]() , ainda mais baixo: 63,2%. A consistência de
PGM difere entre sobreiro (88,8%) e azinheira (79,0%), devendo talvez a priori assumir-se o valor desta
espécie como aplicável aos híbridos e suas descendências.
, ainda mais baixo: 63,2%. A consistência de
PGM difere entre sobreiro (88,8%) e azinheira (79,0%), devendo talvez a priori assumir-se o valor desta
espécie como aplicável aos híbridos e suas descendências.
D — Descendências dos híbridos
Consideram-se separadamente os resultados obtidos com plantas criadas em viveiro a partir de sementes produzidas pelos híbridos SES, SM1 e SM2, onde a dimensão da amostragem pode considerar-se mais ou menos satisfatória, e os obtidos em sementes colhidas em 2000 destes híbridos e ainda de CAB, MRM e VR, ainda muito preliminares pelo reduzido número de plantas e pela interpretação dos zimogramas ser menos segura.
Consideraram-se primariamente os zimogramas DIA, MDH (anódico), MDH (catódico), GsR e PGM, que formavam o conjunto com maior sensibilidade na tarefa de discriminar entre sobreiro, azinheira e híbrido, recorrendo-se ainda às actividades PGI, EST e PGD quando estas pudessem trazer um esclarecimento suplementar. A amostra 38.29 (de SES) não deu qualquer zimograma, possivelmente por ter havido degradação dos enzimas, e das outras 104 amostras apenas 3 de SM1 foram consideradas contaminantes por serem indistinguíveis de sobreiro (38.16) ou de azinheira (33.4 e 38.21). Mas não se pode excluir que sejam na verdade material introgredido, e assim deduz-se que a metodologia utilizada teve cerca de 97% de sensibilidade para a detecção do fenótipo de descendentes dos híbridos. É um valor muito alto, com tão poucos marcadores diagnosticantes.
Das 101 plantas que se passa a considerar, todas de alguma maneira sugeriam a sua origem híbrida, porque pelo menos um dos marcadores era como nos híbridos. No caso das actividades MDH anódicas, houve casos que apresentaram padrões de bandas desconhecidos quer nas espécies de referência, quer nos híbridos, o que sugeria serem novos recombinantes (Az^ e X^ cf. parte II, secção B1).
Foi possível determinar a origem do pólen para a maior parte destas plantas: as que apresentavam um ou mais marcadores de sobreiro iria indicar que o pólen que lhe dera origem era dessa espécie, e do mesmo modo para pólen de azinheira caso houvesse um ou mais marcadores desta espécie; mas já quando se encontram no mesmo indivíduo marcadores de sobreiro e de azinheira, a única hipótese a considerar será que o pólen era recombinante, provavelmente do próprio híbrido (autopolinização); quando todos os marcadores presentes eram como a árvore que lhes deu origem, ou quando os zimogramas PGM implicavam dúvidas sobre a origem do pólen, esta ficou indeterminada. Na tabela 4.17 exemplifica-se o processo de determinação da origem do pólen, e na tabela 4.18 faz-se um resumo dos resultados obtidos.
Tabela 4.17 — Exemplos de plantas descendentes de híbridos de sobreiro e azinheira, com a dedução da origem provável do pólen que deu origem a cada uma, com base nos seus fenótipos isoenzimáticos (sublinhados os mais importantes para a determinação da origem do pólen). Na 3ª coluna inserem-se os zimogramas PGM e PGD das árvores. Identificação da origem do pólen (última coluna): S — sobreiro; A — azinheira; X — recombinante; ? — indeterminado.
Amostra |
Árvore |
PGM, PGD |
DIA |
MDH+ |
MDH– |
GSR |
PGM |
PGD |
PGI |
EST |
pólen |
38.26 |
SES |
?, 12 |
Sb |
Sb |
|
|
d |
12 |
+ |
12 |
S |
33.16 |
SM1 |
(ad), 12+b |
Sb |
X' |
Sb |
Sb |
ad |
|
Az |
2 |
S |
34.30 |
SM2 |
d, 12 |
Sb |
Sb |
Sb |
|
d |
|
Az |
2 |
S |
36.13 |
SM2 |
d, 12 |
Sb |
X' |
|
Sb |
d |
|
Az |
2 |
S |
36.4 |
SM2 |
d, 12 |
X |
X' |
|
X' |
d |
|
+ |
2 |
S |
33.5 |
SM1 |
(ad), 12+b |
Az |
X |
|
Az |
bd |
|
+ |
12 |
A |
34.26 |
SM2 |
d, 12 |
Az |
X' |
X |
|
|
|
+ |
12 |
A |
38.27 |
SES |
?, 12 |
Az |
Sb |
Sb |
|
cd |
12 |
+ |
12 |
X |
38.3 |
SM1 |
(ad), 12+b |
X |
Sb |
Sb |
Az |
d |
|
+ |
12 |
X |
38.8 |
SM1 |
(ad), 12+b |
X |
X |
|
|
ad |
1+2b |
Az |
23 |
? |
34.23 |
SM2 |
d, 12 |
X |
X' |
X |
|
|
|
+ |
2 |
? |
Tabela 4.18 — Origem provável do pólen que interveio nas descendências (plantas) dos híbridos SES, SM1 e SM2. A percentagem de pólen de sobreiro é referente ao total, sem incluir os indeterminados.
pólen |
SES |
SM1 |
SM2 |
todos |
sobreiro |
11 |
15 |
33 |
59 |
azinheira |
0 |
18 |
5 |
23 |
recombinante |
6 |
2 |
2 |
10 |
indeterminado |
1 |
3 |
5 |
9 |
% sobreiro |
65% |
43% |
83% |
64% |
Os 4 casos indeterminados pela aparente contradição entre PGM e os outros marcadores foram a amostra 38.14 de SM1 (assumindo-se este híbrido como ad) e as amostras 34.20, 34.24 e 34.27 de SM2. Tendo em conta a presumível tendência para a actividade MDH catódica aparentar “Sb” quando na realidade será “X” (conforme se documenta na parte II secção A1, e é discutido no próximo parágrafo), pode admitir-se que em 38.14 e 34.20 o pólen fosse de azinheira. Em qualquer caso, as estimativas da última linha da tabela 4.18 não seriam alteradas substancialmente. Apenas a de SM2 se desvia significativamente de 50% (χ2 = 11,3, 1 g.l., P = 8 × 10–5), mas em SES a ausência de pólen de azinheira (aliás a espécie predominante na vizinhança desta árvore!, cf. “Materiais e métodos” parte I secção C) contrasta com o que se esperaria caso este híbrido tivesse igual receptividade ao pólen das duas espécies; no entanto, não parece tratar-se duma diferença de receptividade, pelo menos a avaliar pelos resultados com sementes de 2000 (cf. secção 2).
Considerando a origem provável do pólen de que descende cada planta, é possível determinar o
genótipo do seu complemento feminino, isto é, do gâmeta recombinante produzido pelo híbrido. O
principal interesse desta determinação é verificar a predição de que, para cada locus, metade dos gâmetas
femininos produzidos pelos híbridos foi portador do alelo de sobreiro, e a outra metade do alelo de
azinheira. Enquanto para DIA, MDH (anódico) e PGM
![]() a percentagem de gâmetas portadores de alelo
de sobreiro estava muito próxima da expectativa (48–52%), para GsR era ligeiramente inferior mas não
significativa (39%; teste χ2, 1 g.l.: P = 29,6%), mas havia um desvio altamente significativo para MDH–
(65%; P = 0,77%).
a percentagem de gâmetas portadores de alelo
de sobreiro estava muito próxima da expectativa (48–52%), para GsR era ligeiramente inferior mas não
significativa (39%; teste χ2, 1 g.l.: P = 29,6%), mas havia um desvio altamente significativo para MDH–
(65%; P = 0,77%).
É provável que o desvio em MDH– se relacione com problemas de visualização, em que um zimograma “X” seja interpretado como “Sb” (cf. parte II, secção A1); sete casos são dignos de nota pela implicação que neles tem esta hipótese: nas amostras 7AB, 33.2, 33.9, 33.10 e 38.5, porque a determinação da origem do pólen se decidiu em função dos zimogramas MDH–, e nas amostras 38.14 e 34.20 porque os respectivos zimogramas PGM eram contraditórios com o genótipo da respectiva árvore-mãe, como já referido acima junto com as amostras 34.24 e 34.27. Porém, efectuando a substituição de “Sb” por “X” nesses 7 casos, o desvio continua a ser significativo (64%; P = 1,01%).
Um teste complementar baseia-se na expectativa de 50% de zimogramas híbridos nos descendentes
em cada um dos loci com genótipo híbrido em SES, SM1 e SM2, independente das frequências de pólen
“Sb”, “Az” ou “X”. Descontando-se uma minoria de casos duvidosos, observaram-se 47 em DIA
(esperados 49), 47 em MDH+ (incluindo as variantes X, X^, X’ e X^’, esperados 48,5), 17 em MDH–
(esperados 30), 18 em GsR (X e X’, esperados 18) e 25 em PGM (ad, bd e cd, apenas os descendentes
de SES e SM1, esperados 22,5); mais uma vez, apenas o caso de MDH– se desviou significativamente
da expectativa (
![]() = 10,42, P = 0,125%), que confirma a interpretação dum défice de visualização da
banda “Az” deste marcador.
= 10,42, P = 0,125%), que confirma a interpretação dum défice de visualização da
banda “Az” deste marcador.
Antes ou durante a maceração das folhas destas plantas verificou-se que algumas (8 descendentes de SM1 e 3 de SM2) tinham características das folhas de azinheira (maior consistência ou textura mais fibrosa); e houve pelo menos 2 (de SM1) cujo contorno espinulado das folhas as assemelhavam às folhas de azinheira, embora não na consistência ou na textura na maceração. Em todos estes casos o pólen que lhes deu origem terá sido, com base nos fenótipos isoenzimáticos que apresentaram, de azinheira (perto de metade, considerando as estimativas da tabela 4.18).
O exame dos haplótipos supostos para os loci codificando DIA, MDH+ e PGM, com vista a determinar se haveria ligação cromossómica entre eles, não foi considerado conclusivo.
O número médio de marcadores híbridos apresentados por cada amostra aumentou com o número de zimogramas específicos revelados, numa proporção média próxima de ½. Verificou-se que a situação observada em SM2, onde apenas 4 dos 5 marcadores revelados eram de híbrido, ocorreu também em dois dos descendentes de SM1, indo assim ao encontro da suposição feita anteriormente de que SM2 não é um descendente directo de sobreiro com azinheira.
Consideraram-se separadamente os zimogramas DIA, MDH (anódico) e GDH, embora com sérias
reservas quanto à fiabilidade de DIA (não exibiu “X” nas descendências dos híbridos
![]() ) e quanto à
classificação em padrões de sobreiro (banda S) azinheira (banda F) e híbrido (bandas F e S) de GDH.
A determinação da origem dos grãos de pólen foi feita para 6 híbridos (SES, SM1, SM2, CAB, MRM,
e VR), mas apenas com 6 sementes de cada um (tabela 4.19).
) e quanto à
classificação em padrões de sobreiro (banda S) azinheira (banda F) e híbrido (bandas F e S) de GDH.
A determinação da origem dos grãos de pólen foi feita para 6 híbridos (SES, SM1, SM2, CAB, MRM,
e VR), mas apenas com 6 sementes de cada um (tabela 4.19).
Tabela 4.19 — Origem presumível do pólen que interveio nas descendências (sementes) dos híbridos. Nas amostras onde o aparecimento dum zimograma (DIA ou MDH) de sobreiro e do zimograma “F” em GDH (que se supõe provisoriamente ser de azinheira), a classificação do pólen como sendo recombinante é feita sob reserva, daí ser apresentada separadamente, como “X?”.
pólen |
CAB |
MRM |
SES |
SM1 |
SM2 |
VR |
total |
sobreiro |
1 |
2 |
|
3 |
1 |
3 |
10 |
azinheira |
3 |
1 |
6 |
1 |
1 |
1 |
13 |
recombinante |
1 |
|
|
1 |
2 |
2 |
6 |
X? |
1 |
3 |
|
1 |
2 |
|
7 |
As dúvidas ainda existentes sobre a hereditariedade dos marcadores GDH leva a que se considere a identificação de origem “X?” como duvidosa; porém, se se confirmar que é de facto pólen “X”, a taxa de autopolinização dos híbridos, detectada neste ponto do ciclo de vida, é bastante elevada (6+7 = 13 num total de 36, isto é, 36% do total).
No que respeita à proporção estimada de gâmetas femininos dos híbridos portadores do alelo de sobreiro em cada locus suposto, foi de 68% em DIA e 58% em MDH (ambos sem desvio significativo da expectativa de 50%), mas de 23% em GDH (χ2 = 7,5, 1 g.l., P = 0,6%).
É ainda de notar em SES uma tendência marcada para pólen de azinheira, quando a nível de plantas jovens, na frutificação de 1998, nenhum caso foi detectado (tabela 4.18).
Quanto à ausência de zimogramas “X” em DIA, é possível que apenas um dos alelos esteja a ser expresso nas sementes, e no caso de ser consistente em termos de origem (materna ou paterna) pode tratar-se do fenómeno epigenético de imprinting [Alleman & Doctor 2000]. Na hipótese de ser sempre o alelo materno para DIA que é expresso nestas sementes, procurou-se uma redefinição da origem do pólen com base nos zimogramas MDH e GDH apenas, mas sem progressos em relação ao que é apresentado na tabela 4.19. É fundamental, para esclarecer estas questões em sementes, que a variação interespecífica para PGM e GsR seja clarificada.
E — Frequências intra-específicas
1) Povoamentos mistos (adultos) e de referência (plantas jovens)
Na tabela 4.20 apresentam-se as frequências dos 5 alelos identificados no presente estudo nas azinheiras das populações de referência (plantas jovens) e montados mistos (adultos).
Tabela 4.20 — Frequências dos alelos no locus Pgm de azinheira, a partir da amostragem de adultos nos povoamentos mistos e de plantas jovens nos povoamentos de referência, com arredondamento à 4ª casa decimal. N, número de zimogramas observados.
|
Banda |
a' |
a |
b |
c |
d |
|
Pgm |
Alelo |
1 |
2 |
3 |
4 |
5 |
N |
mistos |
Mitra |
0 |
0,1 |
0,16 |
0,74 |
0 |
25 |
Feijoas |
0,025 |
0,175 |
0,2 |
0,575 |
0,025 |
20 |
|
Alfaiates |
0 |
0,1667 |
0,2 |
0,6333 |
0 |
15 |
|
Conjunto |
0,0083 |
0,1417 |
0,1833 |
0,6583 |
0,0083 |
60 |
|
referência |
Azeiteiros |
0 |
0,1293 |
0,3146 |
0,5463 |
0,0098 |
205 |
Testa |
0,0073 |
0,0985 |
0,2007 |
0,6679 |
0,0255 |
137 |
A análise das distâncias genéticas de Nei, neste locus, sugere que as azinheiras de Azeiteiros se diferenciam mais do que as dos povoamentos mistos e de Testa (entre Feijoas e Alfaiates com um valor bastante baixo D = 0,0025). O valor de GST entre os 3 povoamentos mistos situou-se em 1,23%, sugerindo uma baixa diferenciação populacional, corroborada pelo teste através duma tabela de contingência, que deu χ2 = 3,42 (4 g.l. juntando os alelos 1, 2 e 5 numa só classe, P = 0,49), donde se conclui que as frequências neste locus são homogéneas entre os 3 povoamentos mistos, deste modo validando a utilização das frequências com base no conjunto de 60 observações. O programa GENEPOP confirmou esta conclusão (diferenciação génica, P = 0,65; diferenciação genotípica, P = 0,77).
Na tabela 4.21 resumem-se os parâmetros de Gregorius para este locus. Os valores parecem bastante homogéneos entre povoamentos, embora sejam patentes valores mais baixos de ν e δT na Mitra, e um Dj mais elevado na Mitra e em Azeiteiros.
Tabela 4.21 — Estatísticas populacionais de Gregorius (diversidade genética ou número efectivo de alelos ν, diferenciação total no povoamento δT, diferenciação de cada povoamento em relação aos restantes Dj) no locus Pgm de azinheira, a partir dos valores das frequências na tabela 4.20.
Parâmetro |
Mitra |
Feijoas |
Alfaiates |
Conjunto |
Azeiteiros |
Testa |
ν |
1,715 |
2,484 |
2,133 |
2,052 |
2,414 |
2,013 |
δT |
0,434 |
0,629 |
0,569 |
0,521 |
0,589 |
0,507 |
Dj |
0,134 |
0,091 |
0,042 |
|
0,124 |
0,062 |
Dj |
|
|
|
0,084 |
0,132 |
0,085 |
As distribuições genotípicas nos povoamentos mistos não se afastam significativamente das esperadas no modelo de Hardy-Weinberg, conforme documenta a tabela 4.22.
Tabela 4.22 — a) Distribuição dos fenótipos PGM de azinheira nos povoamentos mistos, separadamente e em conjunto, e teste χ2 (1 g.l., com correcção de Yates) para os valores esperados (distribuição de Hardy-Weinberg) do fenótipo c, heterozigóticos (_c) com a banda c, e restantes (etc), representados com 4 algarismos significativos. b) Teste χ2 (1 g.l.) para os valores esperados (distribuição de Hardy-Weinberg) de 5 classes fenotípicas no conjunto dos povoamentos. Sublinham-se, em ambos os testes, os valores esperados abaixo de 5.
a) |
PGM |
|
valores esperados |
|
|
|||||||||
|
a |
ab |
ac |
a'c |
b |
bc |
bd |
c |
total |
c |
_c |
etc |
χ2 |
P |
Mitra |
|
1 |
4 |
|
1 |
5 |
|
14 |
25 |
13,69 |
9,620 |
1,690 |
0,03 |
0,87 |
Feijoas |
2 |
1 |
2 |
1 |
1 |
4 |
1 |
8 |
20 |
6,612 |
9,775 |
3,612 |
0,87 |
0,35 |
Alfaiates |
1 |
|
3 |
|
2 |
2 |
|
7 |
15 |
6,017 |
6,967 |
2,017 |
0,46 |
0,50 |
Conjunto |
3 |
2 |
9 |
1 |
4 |
11 |
1 |
29 |
60 |
26,00 |
26,99 |
7,004 |
2,25 |
0,13 |
b) |
valores esperados |
|
|
||||
|
ac |
ab+b |
bc |
c |
etc |
χ2 |
P |
Conjunto |
11,19 |
5,133 |
14,48 |
26,00 |
3,188 |
2,79 |
0,09 |
Os valores do coeficiente de fixação foram de 0,04 em Mitra, 0,25 em Feijoas e 0,37 em Alfaiates, mas trata-se de valores talvez pouco representativos pela reduzida dimensão das amostragens respectivas (aliás, o valor do FST resultante foi negativo), enquanto no conjunto se obteve 0,22.
Nos povoamentos de referência, só se observaram desvios significativos em relação à expectativa do modelo de Hardy-Weinberg em Azeiteiros, principalmente pelo déficit de indivíduos ab (tabela 4.23). Este desvio pode ser fortuito, por isso, apesar do erro de tipo I ser pequeno (0,035), pode não ser correcto rejeitar a hipótese de conformidade com o modelo (aliás, o valor de P = 0,25 obtido com o programa GENEPOP parece confirmar esta interpretação).
Tabela 4.23 — Distribuição dos fenótipos PGM de azinheira nos povoamentos de referência (plantas jovens), e teste χ2 (1 g.l.) para os valores esperados (distribuição de Hardy-Weinberg), representados com 4 algarismos significativos (d_ representa todos as classes fenotípicas com banda d, e a'_ as com a banda a', excepto a'd).
|
PGM |
|
|
||||||||||
|
a'c |
a'd |
a |
ab |
ac |
ad |
b |
bc |
bd |
c |
cd |
total |
F |
Azeiteiros |
|
|
6 |
10 |
31 |
|
24 |
70 |
1 |
60 |
3 |
205 |
0,04 |
Testa |
1 |
1 |
2 |
7 |
15 |
1 |
6 |
34 |
2 |
65 |
3 |
137 |
0,07 |
|
valores esperados |
|
||||||
|
ab |
ac |
b |
bc |
c |
d_+a+a'_ |
χ2 |
P |
Azeiteiros |
16,68 |
28,96 |
20,29 |
70,48 |
61,19 |
7,406 |
4,43 |
0,04 |
Testa |
5,420 |
18,03 |
5,520 |
36,73 |
61,11 |
10,18 |
1,53 |
0,22 |
Em conformidade com a nomenclatura adoptada por outros autores [Jiménez et al. 1999], o locus para as actividades polimórficas foi designado Pgi-B. Na tabela 4.24 apresentam-se as frequências dos 3 alelos identificados no presente estudo nos sobreiros da população de referência (plantas jovens) e montados mistos (adultos).
Tabela 4.24 — Frequências dos alelos no locus Pgi-B de azinheira, a partir da amostragem de adultos nos povoamentos mistos e de plantas jovens nos povoamentos de referência, com arredondamento à 4ª casa decimal. N, número de zimogramas observados.
|
Banda |
a |
b |
c |
|
Pgi-B |
Alelo |
1 |
2 |
3 |
N |
mistos |
Mitra |
0,5875 |
0,3875 |
0,025 |
40 |
Feijoas |
0,7419 |
0,2581 |
0 |
31 |
|
Alfaiates |
0,725 |
0,275 |
0 |
20 |
|
Conjunto |
0,6703 |
0,3187 |
0,011 |
91 |
|
referência |
Me. Branco |
0,5763 |
0,4237 |
0 |
177 |
A forte semelhança entre as frequências dos alelos predominantes (Pgi-B1 e Pgi-B2) e as obtidas de populações da Península Ibérica noutros estudos [Toumi & Lumaret 1998, Jiménez 2001], a que acresce a identidade das taxas de migração implícitas na designação dos alelos por Toumi & Lumaret [1998] (Pgi1.90 o mais frequente, Pgi1.70 o segundo, e Pgi1.50 ou terceiro, correspondendo às migrações tipificadas pelas posições a, b e c no presente estudo, tabela 4.8) sugerem tratar-se do mesmo locus.
A análise das distâncias genéticas de Nei, neste locus, sugere uma maior identidade entre os sobreiros da Mitra e os de Monte Branco (D = 0,0019), e entre os de Feijoas e Alfaiates (esta com um valor extremamente baixo D = 0,0004). Mesmo assim, o valor de GST entre os 3 povoamentos mistos situou-se em 1,88%, sugerindo uma baixa diferenciação populacional, corroborada pelo teste através duma tabela de contingência, que deu χ2 = 4,46 (2 g.l. juntando os alelos 2 e 3 numa só classe, P = 0,11), donde se conclui que as frequências neste locus são homogéneas entre os 3 povoamentos mistos, deste modo validando a utilização das frequências com base no conjunto de 91 observações. O programa GENEPOP confirmou esta conclusão (diferenciação génica, P = 0,19; diferenciação genotípica, P = 0,13), mas não a presumida proximidade entre Mitra e Monte Branco.
Na tabela 4.25 resumem-se os parâmetros de Gregorius para este locus. Os valores parecem bastante homogéneos entre povoamentos, embora seja patente um valor mais elevado de Dj em Feijoas e em Monte Branco.
Tabela 4.25 — Estatísticas populacionais de Gregorius no locus Pgi-B de sobreiro, a partir dos valores das frequências na tabela 4.24.
Parâmetro |
Mitra |
Feijoas |
Alfaiates |
Conjunto |
Me. Branco |
ν |
2,016 |
1,621 |
1,663 |
1,815 |
1,955 |
δT |
0,517 |
0,396 |
0,420 |
0,454 |
0,491 |
Dj |
0,094 |
0,112 |
0,090 |
|
0,117 |
As distribuições genotípicas nos povoamentos mistos não se afastam significativamente das esperadas no modelo de Hardy-Weinberg, conforme documenta a tabela 4.26.
Tabela 4.26 — Distribuição dos fenótipos PGI de sobreiro nos povoamentos mistos, separadamente e em conjunto, e teste χ2 (1 g.l., com correcção de Yates) para os valores esperados (distribuição de Hardy-Weinberg) do fenótipo a, heterozigóticos (_a) com a banda a, e restantes (etc), representados com 4 algarismos significativos. Sublinham-se os valores esperados abaixo de 5.
|
PGI |
|
valores esperados |
|
|
|||||
|
a |
ab |
b |
bc |
total |
a |
ab |
b+bc |
χ2 |
P |
Mitra |
16 |
15 |
7 |
2 |
40 |
13,81 |
19,39 |
6,806 |
1,46 |
0,23 |
Feijoas |
16 |
14 |
1 |
|
31 |
17,06 |
11,87 |
2,065 |
0,52 |
0,47 |
Alfaiates |
9 |
11 |
|
|
20 |
10,51 |
7,975 |
1,513 |
1,86 |
0,17 |
Conjunto |
41 |
40 |
8 |
2 |
91 |
40,89 |
40,22 |
9,890 |
0,02 |
0,88 |
Os valores do coeficiente de fixação foram de 0,16 em Mitra, –0,18 em Feijoas e –0,38 em Alfaiates, mas trata-se de valores talvez pouco representativos pela reduzida dimensão das amostragens respectivas (o GENEPOP dá estimativas muito semelhantes, com valores de P não significativos); no conjunto obteve-se uma estimativa de –0,03, com um FST de 4,5% (1.7% usando a definição de Weir & Cockerham [1984], calculada por GENEPOP).
No povoamento de referência, porém, observaram-se desvios significativos em relação à expectativa do modelo de Hardy-Weinberg (tabela 4.27), relacionadas com um excedente de homozigóticos traduzido num valor F = 0,33.
Tabela 4.27 — Distribuição dos fenótipos PGI de sobreiro no povoamento de referência de Monte Branco (plantas jovens), e teste χ2 (1 g.l.) para os valores esperados (distribuição de Hardy-Weinberg), representados com 4 algarismos significativos.
|
PGI |
|
valores esperados |
|
|
||||
|
a |
ab |
b |
total |
a |
ab |
b |
χ2 |
P |
Monte Branco |
73 |
58 |
46 |
177 |
58,78 |
86,44 |
31,78 |
19,16 |
< 0,001 |
O conhecimento, para os povoamentos mistos, dos genótipos das plantas e das suas progenitoras, permitiu estimar as frequências alélicas na fase haplóide (dos gâmetas femininos e masculinos que deram origem a essas plantas), compará-los com os das plantas, e calcular o coeficiente de fixação. Depois de excluídos os contaminantes (cf. tabela 4.10 para o locus Pgm), em cada família procedeu-se à comparação entre os zimogramas da árvore e das descendentes, em geral permitindo identificar o alelo transmitido pelo pólen — por exemplo, 5 descendentes da árvore 27 de Alfaiates (cujo zimograma de PGI é a) apresentaram zimograma PGI a, e 1 o zimograma ab, donde se deduz que houve 5 grãos de pólen portadores do alelo Pgi-B1, e 1 do alelo Pgi-B2. Noutro exemplo, à árvore 60 da Mitra, com zimograma PGM bc, corresponderam 45 plantas distribuídas por 6 classes de zimogramas diferentes (2 com zimograma ab, 2 b, 1 a'c, 7 ac, 24 c, e 9 bc); considerando os 5 alelos do locus Pgm, contar-se-iam para as primeiras 5 classes outros tantos grãos de pólen portadores dos alelos Pgm2, Pgm3, Pgm1, Pgm2, Pgm4, respectivamente, mas para a classe bc poderiam ser portadores de Pgm3 ou Pgm4 — embora estes 9 casos formalmente fossem indeterminados, teoricamente 4,5 seriam portadores de Pgm3 e os outros 4,5 de Pgm4. As tabelas 4.28 e 4.29 resumem a informação obtida para os dois loci.
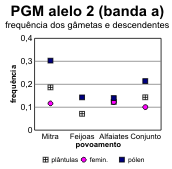
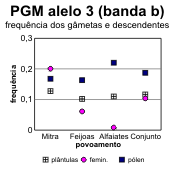
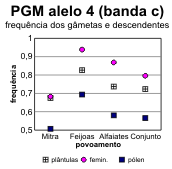
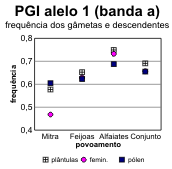
A partir destes dados pode fazer-se o cálculo das frequências alélicas em cada locus, tanto para os dois tipos de gâmetas na fase haplóide como para as plantas (tabelas 4.30 e 4.31) [Bacilieri et al. 1996]. Nos gráficos da figura 4.17 faz-se a correspondente ilustração. É patente a variação nas frequências entre os três povoamentos, o que deverá reflectir um efeito da amostragem, e a par disso uma discrepância quase generalizada entre gâmetas femininos e masculinos, no que respeita às respectivas frequências. Onde o desvio entre as frequências nos gâmetas são mais pronunciados (principalmente no caso do locus Pgm), as frequências nas plantas situam-se em geral num valor intermédio entre as dos gâmetas (excepção apenas para o alelo 3 na Mitra).
Fazendo o cálculo de F = 1 – Ho/He para as plantas (tabela 4.32), resulta para o conjunto dos povoamentos mistos um valor de –0,049 para o locus Pgm de azinheira e de 0,074 para o locus Pgi-B de sobreiro. No entanto, estes valores só têm significado assumindo que as frequências são homogéneas entre gâmetas masculinos e femininos. Não o sendo, como sugerem as tabelas 4.30 e 4.31 (e figura 4.17), os valores de He serão diferentes (modelo II na tabela 4.32), de que resulta F = 0,028 e 0,080, respectivamente. Mais ainda, os valores de referência para estes F nas plantas, reflectindo a discrepância (correlação negativa) entre os gâmetas, é negativo, o que pode significar uma estimativa de Ne bastante baixa, na ordem dos 4,66 para azinheira e 5,87 para sobreiro.
Em referência ao modelo de Wright de subdivisão populacional, os valores de Fobs na tabela 4.32 são o FIT, e a partir do He médio (entre povoamentos) obtêm-se no modelo I FIS = –0,138 para Pgm (azinheira) e 0,102 para Pgi-B (sobreiro); mas se no primeiro caso o FST resultante é 0,078, no segundo é negativo. Usando os He do modelo II, tem-se FIS = –0,046 (FST = 0,070) para Pgm e FIS = 0,108 (FST negativo) para Pgi-B. Dado tratar-se apenas de 3 subpopulações, é de admitir que os dados sejam pouco adequados para este tipo de análise. Com estes valores de FST para Pgm deduz-se Nem = 1,32 ou 1,47 (modelo I ou modelo II, respectivamente). Da estimativa de Ne dada acima para o modelo II (4,66) obtém-se m = 0,315.
Pode estimar-se a percentagem de pólen não-aparentado geneticamente, definida como t = (1 – F)/(1 + F) [Brown & Allard 1970], mas substituindo F pelo valor ΔF, visto que o valor de referência para F não é 0. Assim, tPgm = 81% e tPgi-B = 84%.
Todos os resultados em Pgm de azinheira se mantêm, no essencial, caso se considere que a árvore A21 tinha genótipo ac, deste modo integrando-se um total de 27 plantas (cf. tabela 4.10 e texto respectivo); por exemplo, a estimativa de Ne é 4,51, a de m 0,346, e a de tPgm 80%.
Tabela 4.28 — Frequências para os zimogramas PGM nas plantas e frequências para os alelos do locus Pgm de azinheira nos gâmetas que lhes deram origem, referentes aos povoamentos mistos.
|
|
gâmetas femininos |
zimogramas PGM |
gâmetas masculinos |
||||||||||||||
|
plantas |
Pgm2 |
Pgm3 |
Pgm4 |
a'a |
a |
ab |
b |
a'c |
ac |
bc |
c |
cd |
Pgm1 |
Pgm2 |
Pgm3 |
Pgm4 |
Pgm5 |
Mitra |
137 |
16 |
27,5 |
93,5 |
|
4 |
4 |
3 |
3 |
39 |
25 |
59 |
|
3 |
41,5 |
23 |
69,5 |
|
Feijoas |
49 |
0 |
3 |
46 |
|
|
|
1 |
|
7 |
8 |
33 |
|
|
7 |
8 |
34 |
|
Alfaiates |
118 |
14,5 |
1 |
102,5 |
1 |
2 |
2 |
|
3 |
22 |
24 |
61 |
3 |
4 |
16,5 |
26 |
68,5 |
3 |
Conjunto |
304 |
30,5 |
31,5 |
242 |
1 |
6 |
6 |
4 |
6 |
68 |
57 |
153 |
3 |
7 |
65 |
57 |
172 |
3 |
Tabela 4.29 — Frequências para os zimogramas PGI (polimórfica) nas plantas, e frequências para os alelos do locus Pgi-B de sobreiro nos gâmetas que lhes deram origem, referentes aos povoamentos mistos.
|
|
gâmetas femininos |
zimogramas PGI |
gâmetas masculinos |
||||||
|
plantas |
Pgi-B1 |
Pgi-B2 |
a |
ab |
b |
ac, bc |
Pgi-B1 |
Pgi-B2 |
Pgi-B3 |
Mitra |
110 |
51,5 |
58,5 |
37 |
53 |
19 |
1 |
66,5 |
42,5 |
1 |
Feijoas |
167 |
105 |
62 |
75 |
68 |
23 |
1 |
104 |
62 |
1 |
Alfaiates |
333 |
244 |
89 |
190 |
118 |
24 |
1 |
229 |
103 |
1 |
Conjunto |
610 |
400,5 |
209,5 |
302 |
239 |
66 |
3 |
399,5 |
207,5 |
3 |
Tabela 4.30 — Frequências dos alelos no locus Pgm de azinheira, considerando os gâmetas femininos, gâmetas masculinos e plantas.
alelo |
2 |
3 |
4 |
1 e 5 |
||||||||||||
pov. |
Mitra |
Feij. |
Alf. |
todos |
Mitra |
Feij. |
Alf. |
todos |
Mitra |
Feij. |
Alf. |
todos |
Mitra |
Feij. |
Alf. |
todos |
Femin. |
0,117 |
0,000 |
0,123 |
0,100 |
0,201 |
0,061 |
0,008 |
0,104 |
0,682 |
0,939 |
0,869 |
0,796 |
0,000 |
0,000 |
0,000 |
0,000 |
Masc. |
0,303 |
0,143 |
0,140 |
0,214 |
0,168 |
0,163 |
0,220 |
0,188 |
0,507 |
0,694 |
0,581 |
0,566 |
0,022 |
0,000 |
0,059 |
0,033 |
Plant. |
0,186 |
0,071 |
0,123 |
0,143 |
0,128 |
0,102 |
0,110 |
0,117 |
0,675 |
0,827 |
0,737 |
0,724 |
0,011 |
0,000 |
0,030 |
0,016 |
Tabela 4.31 — Frequências dos alelos no locus Pgi-B de sobreiro, considerando os gâmetas femininos, gâmetas masculinos e plantas.
alelo |
1 |
2 |
3 |
|||||||||
pov. |
Mitra |
Feij. |
Alf. |
todos |
Mitra |
Feij. |
Alf. |
todos |
Mitra |
Feij. |
Alf. |
todos |
Femin. |
0,468 |
0,629 |
0,733 |
0,657 |
0,532 |
0,371 |
0,267 |
0,343 |
0,000 |
0,000 |
0,000 |
0,000 |
Masc. |
0,605 |
0,623 |
0,688 |
0,655 |
0,386 |
0,371 |
0,309 |
0,340 |
0,009 |
0,006 |
0,003 |
0,005 |
Plant. |
0,577 |
0,653 |
0,749 |
0,691 |
0,418 |
0,344 |
0,249 |
0,307 |
0,005 |
0,003 |
0,002 |
0,002 |
a)
 b)
b)
 c)
c)
d)
 e)
e)
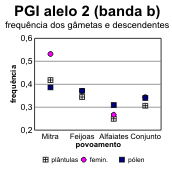
Figura 4.17 — Frequências dos alelos comuns no locus Pgm de azinheira (gráficos a, b, c) e no locus Pgi-B de sobreiro (gráficos d, e), estimadas para os dois tipos de gâmetas e para as plantas nos povoamentos mistos.
Tabela 4.32 — Valores observados (Ho) e esperados (He) de frequência de heterozigóticos, calculados levando em conta as frequências alélicas nas plantas (modelo I) ou levando em conta os desvios Δxi entre essas frequências e as estimadas para o pólen (tabelas 4.30 e 4.31, modelo II). O coeficiente de fixação Fobs = 1–Ho/He resultante tem por referência o valor 0 no modelo I e o valor Fesp no modelo II; por isso, no modelo II calcula-se o Ne em função da diferença Fobs–Fesp (ΔF).
|
Locus |
Pgm (azinheira) |
Pgi-B (sobreiro) |
||||||||
|
|
Mitra |
Feij. |
Alf. |
média |
todos |
Mitra |
Feij. |
Alf. |
média |
todos |
|
Ho |
0,518 |
0,306 |
0,466 |
|
0,464 |
0,491 |
0,413 |
0,357 |
|
0,397 |
I |
He |
0,493 |
0,301 |
0,429 |
0,408 |
0,442 |
0,492 |
0,455 |
0,378 |
0,442 |
0,429 |
|
Fobs |
–0,051 |
–0,016 |
–0,087 |
|
–0,049 |
0,002 |
0,093 |
0,055 |
|
0,074 |
|
Ne |
|
|
|
|
|
276,78 |
5,39 |
9,16 |
|
6,73 |
II |
He |
0,537 |
0,328 |
0,466 |
0,444 |
0,477 |
0,494 |
0,457 |
0,389 |
0,445 |
0,431 |
|
Fobs |
0,034 |
0,066 |
0,000 |
|
0,028 |
0,006 |
0,096 |
0,069 |
|
0,080 |
|
Fesp |
–0,088 |
–0,088 |
–0,087 |
|
–0,079 |
–0,004 |
–0,004 |
–0,020 |
|
–0,006 |
|
ΔF |
0,123 |
0,154 |
0,088 |
|
0,107 |
0,009 |
0,100 |
0,089 |
|
0,085 |
|
Ne |
4,08 |
3,25 |
5,71 |
|
4,66 |
54,72 |
5,02 |
5,63 |
|
5,87 |
Das análises de rotina, visando primariamente a validação dos marcadores discriminantes e a detecção de híbridos, foi possível:
1. A confirmação dos contaminantes interspecíficos e a detecção de parte dos intra-específicos.
2. A caracterização parcial de 9 híbridos, sendo que SM2 pode já ser descendente de híbrido por retrocruzamento com sobreiro.
3. Na ausência de híbridos nas descendências dos povoamentos mistos em 1998, o cálculo de majorantes da probabilidade de hibridação, para esses povoamentos e nesse ano, sendo a estimativa mais conservadora de P < 0,043%.
4. A caracterização de adultos e plantas jovens com fenótipos atípicos, sendo que o número de casos de introgressão parece ser muito reduzido.
5. A caracterização de 45 amostras de sobreiros de cortiça preguenta, tendo-se detectado em 2 marcadores (DIA e PER) sinais de possível introgressão de genes de azinheira nalgumas árvores.
6. Uma quantificação dos valores de especificidade e sensibilidade, na distinção de híbridos e azinheiras em relação a sobreiro, dos marcadores utilizados nas análises de rotina.
7. A determinação da origem do pólen nas descendências dos híbridos SM1, SM2 e SES.
8. A análise das frequências intra-específicas nos loci Pgm (azinheiras) e Pgi-B (sobreiros), com relevo para considerarem-se os três povoamentos mistos, como fazendo parte duma mesma população, em ambas espécies, e a determinação de vários estimadores populacionais.
9. A análise familiar para os mesmos loci revelou diferenças entre as distribuições de frequências no pólen, gâmetas femininos e plantas, especialmente em Pgm, e permitiu determinar valores neutrais de Ne na ordem de 5 a 7 em ambas as espécies.
10. A estimativa para a percentagem de pólen não-aparentado é de 81% para azinheira (Pgm) e 84% para sobreiro (Pgi-B).